
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
| Рис.6-6. |
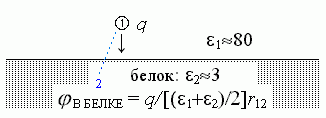
Очень похожий ответ получится, если мы будем рассматривать комплементарную задачу: о поле заряда, находящегося под поверхностью белка (Рис.6-7).
| Рис.6-7. |

В этом случае эффективная диэлектрическая проницаемость eэфф тоже близка к (e1+e2)/2, т. е. к 40 всюду на больших расстояниях от заряда q, а к e2 (т. е. к 3), — только когда r12 много меньше, чем расстояние от заряда q до поверхности белка.
Еще более любопытный результат дает задача о поле, которое заряд, находящийся на одном краю белка, создает на другом краю белка (Рис.6-8).
| Рис.6-8. |
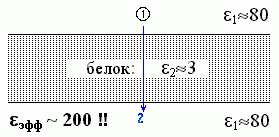
Казалось бы, в этом случае, поскольку взаимодействие идет через белок, через среду с низкой (e2»3) диэлектрической проницаемостью, — мы должны были бы ожидать, что eэфф будет близко к 3, в крайнем случае — ведь вода вокруг — лежать где-то между 3 и 80. А на самом деле — eэфф зашкаливает за 200! Как так??
Для объяснения посмотрим, как ориентируются в этом случае полярные молекулы воды вокруг белка и вокруг заряда ![]() , и как это меняет поле заряда (см. Рис.6-9; он дан, для наглядности, в несколько другом масштабе, чем Рис.6-8, но описывает то же явление).
, и как это меняет поле заряда (см. Рис.6-9; он дан, для наглядности, в несколько другом масштабе, чем Рис.6-8, но описывает то же явление).
Рис.6-9. Ориентация молекул воды (они изображены в виде диполей |
|
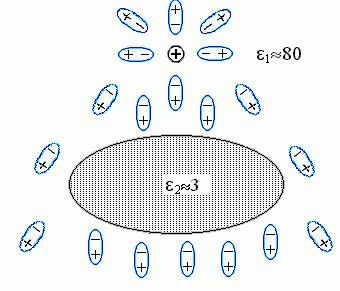
Молекулы воды ориентируются по полю: их "-" преимущественно смотрит в сторону заряда ![]() , а "+" — в противоположную сторону. В результате, во-первых, заряд частично компенсируется примыкающими к нему "минусами" молекул воды; это — тривиальное явление, оно просто приводит к появлению диэлектрической проницаемости воды, e1. Во-вторых — возникает то явление, которое нас интересует. А именно, — повернутые к белку "плюсы" молекул воды, примыкающих к белку с той стороны, где находится заряд
, а "+" — в противоположную сторону. В результате, во-первых, заряд частично компенсируется примыкающими к нему "минусами" молекул воды; это — тривиальное явление, оно просто приводит к появлению диэлектрической проницаемости воды, e1. Во-вторых — возникает то явление, которое нас интересует. А именно, — повернутые к белку "плюсы" молекул воды, примыкающих к белку с той стороны, где находится заряд ![]() , создают на этой стороне поляризационный заряд "+"; а "минусы" молекул воды, примыкающих к белку с другой стороны, создают там противоположный поляризационный заряд "-" (поляризацией самого тела белка можно пренебречь, — по сравнению с поляризацией воды она мала, так как его e2 <<e1).
, создают на этой стороне поляризационный заряд "+"; а "минусы" молекул воды, примыкающих к белку с другой стороны, создают там противоположный поляризационный заряд "-" (поляризацией самого тела белка можно пренебречь, — по сравнению с поляризацией воды она мала, так как его e2 <<e1).
Сумма поляризационных зарядов у поверхности исходно незаряженного тела, окруженного однородной средой (у нас — водой), должна быть равна нулю — есть такая теорема в электростатике. Значит, суммарный поляризационный заряд "+" у поверхности белка обращен к ![]() (!!), а равный ему поляризационный "-" (у другой его поверхности), — в обратную сторону.
(!!), а равный ему поляризационный "-" (у другой его поверхности), — в обратную сторону.
В результате на обращенной к заряду стороне белка потенциал поля возрастает по сравнению с тем, что было бы без белка, — здесь, в дополнение к потенциалу заряда ![]() , потенциал создается и поляризационными "плюсами" у этой поверхности белка (а поляризационные "минусы" от этого места далеко и влияют мало). Поэтому здесь eэфф»40, как о том уже говорилось.
, потенциал создается и поляризационными "плюсами" у этой поверхности белка (а поляризационные "минусы" от этого места далеко и влияют мало). Поэтому здесь eэфф»40, как о том уже говорилось.
В то же время, на отвернутой от заряда стороне белка потенциал поля падает по сравнению с тем, что было бы без белка, — здесь к потенциалу заряда ![]() добавляется противоположный по знаку потенциал поляризационных "минусов" у ближайшей поверхности белка (а поляризационные "плюсы" от этого места далеко и влияют мало). А раз здесь потенциал поля упал по сравнению с тем, что было бы без белка, — то здесь (на противоположной
добавляется противоположный по знаку потенциал поляризационных "минусов" у ближайшей поверхности белка (а поляризационные "плюсы" от этого места далеко и влияют мало). А раз здесь потенциал поля упал по сравнению с тем, что было бы без белка, — то здесь (на противоположной ![]() 'у стороне) eэффбольше, чем то e1, что было без белка, — т. е. больше, чем e1=80.
'у стороне) eэффбольше, чем то e1, что было без белка, — т. е. больше, чем e1=80.
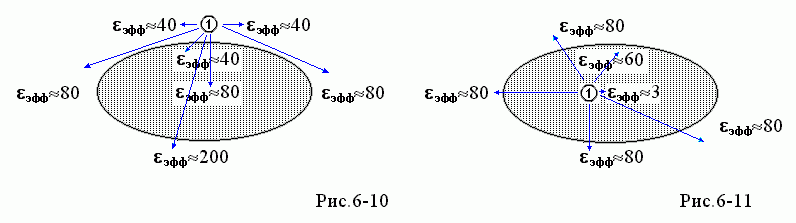
В целом, распределение значений eэфф "в белке и вокруг" для поля, создаваемого зарядом ![]() и наведенного им же поляризационными зарядами, выглядит так, как на Рис.6-10, если заряд
и наведенного им же поляризационными зарядами, выглядит так, как на Рис.6-10, если заряд ![]() находится у поверхности белка, и так, как на Рис.6-11, если заряд
находится у поверхности белка, и так, как на Рис.6-11, если заряд ![]() находится в глубине белка.
находится в глубине белка.
Напоминаю, что eэфф — это та эффективная величина диэлектрической проницаемости для точки r, которую нужно подставлять в формулу расчета потенциала заряда "1" в точке r: f(r)= q1/eэфф|r-r1|.
Отметим еще одно явление, вытекающее из наличия поверхности раздела и связанных с ней "отраженных зарядов". Оно касается действия заряда на самого себя [см. также обсуждение формулы (6.3)]: находящийся вне белка заряд отталкивается от поверхности белка, а заряд, находящийся внутри белка, сильно притягивается к его поверхности — т. е., в обоих случаях, среда с более высокой диэлектрической проницаемостью притягивает заряд, а среда с более низкой — его выталкивает. Силу этого притяжения и выталкивания легко оценить — ведь речь здесь идет о взаимодействии заряда с его отражениями (см. Рис.6-5, 6-6), но в этом я рекомендую вам попрактиковаться самостоятельно.
Теперь займемся эффектами, связанными с корпускулярностью, т. е. атомарным строением среды.
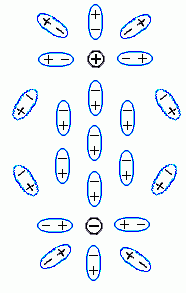
Собственно говоря, сама величина диэлектрической проницаемости e определяется атомарным строением среды. Если среда состоит из неполярных молекул, электрическое поле лишь смещает электроны в них, что сделать трудно: поэтому электроны смещаются мало, и e невелико. Если среда состоит из полярных молекул (пример — вода), электрическое поле разворачивает эти молекулы; это сделать легче, и e такой среды велико. В обоих случаях — и при смещении электронов, и при повороте полярных молекул, — поляризация среды как бы частично "гасит" внесенные в нее заряды (заряды ![]() и
и ![]() , Рис.6-12), и тем самым уменьшает электрическое поле в среде, — по сравнению с тем, что было бы в вакууме.
, Рис.6-12), и тем самым уменьшает электрическое поле в среде, — по сравнению с тем, что было бы в вакууме.
| Рис.6-12. |
Естественно было бы предположить, что связанные с корпускулярностью эффекты должны сильно влиять на взаимодействие зарядов на малых расстояниях — ведь классические формулы (6.1 — 6.3) справедливы, строго говоря, только когда между взаимодействующими зарядами находится много молекул среды. А если заряды (как часто бывает в белках) находятся на расстоянии 3-4 ![]() , никакой другой атом между ними уже не влезет и не изменит их взаимодействия.
, никакой другой атом между ними уже не влезет и не изменит их взаимодействия.
Казалось бы, в случае столь тесного контакта зарядов — диэлектрическая проницаемость для их взаимодействия должна приближаться к 1 даже в водном окружении. Эту точку зрения — точнее, это опасение — до сих пор можно встретить в литературе.
Однако, как ни странно, корпускулярность среды не меняет кардинально "макроскопическую" (т. е. выведенную для больших расстояний между зарядами) диэлектрическую проницаемость среды даже на расстоянии порядка 3 ![]() . То есть даже здесь eэфф гораздо ближе к 80 или 40, чем к 1 или 3.
. То есть даже здесь eэфф гораздо ближе к 80 или 40, чем к 1 или 3.
Об этом свидетельствует то, что соль хорошо растворяется (диссоциирует) в воде, что возможно только при слабом притяжении противоионов даже на самых малых, »3![]() расстояниях.
расстояниях.
В самом деле, ионы Na+ и Cl - могут сблизиться до расстояния 3![]() . При этом свободная энергия их притяжения составляла бы —1.5 ккал/моль при e=80, —3 ккал/моль при e=40, и —6 ккал/моль при e=20. Последнее (6 ккал/моль) превосходит энергию водородной связи. При такой энергии противоионы слипались бы друг с другом сильнее, чем молекулы воды, и тогда насыщенный раствор соли имел бы концентрацию порядка 10-4 моля на литр — как насыщенный водный пар. Но это явно неправда: растворить 1 моль NaCl (58 г) в литре воды — не проблема (это — обычный, может чуть крепкий рассол). Значит, диэлектрическая проницаемость воды существенно больше 20 даже на расстоянии »3
. При этом свободная энергия их притяжения составляла бы —1.5 ккал/моль при e=80, —3 ккал/моль при e=40, и —6 ккал/моль при e=20. Последнее (6 ккал/моль) превосходит энергию водородной связи. При такой энергии противоионы слипались бы друг с другом сильнее, чем молекулы воды, и тогда насыщенный раствор соли имел бы концентрацию порядка 10-4 моля на литр — как насыщенный водный пар. Но это явно неправда: растворить 1 моль NaCl (58 г) в литре воды — не проблема (это — обычный, может чуть крепкий рассол). Значит, диэлектрическая проницаемость воды существенно больше 20 даже на расстоянии »3![]() .
.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
