
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
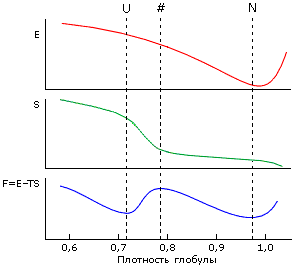
Рис.17-7. Изменение энергии E, энтропии S и свободной энергии F=E-TS с изменением плотности глобулы. D — денатурированное состояние (в данном случае — расплавленная глобула, так как плотность его велика), N — нативная глобула, # — "барьер", максимум свободной энергии на пути равномерного расширения глобулы (от N к D).
Обратимся еще раз к уравнениям (17.1). Они показывают, что отношение скоростей прямой и обратной реакций, kА®В/kВ®А, есть просто константа равновесия между финальным (В) и начальным (А) состояниями
КВ:А = kА®В/kВ®А = exp[-(FВ-FA)/RT] . | (17.4) |
Величина константы равновесия КВ:А есть nB¥/nА¥, она показывает то соотношение между финальным числом nB¥ молекул в состоянии В и их финальным числом nA¥ в состоянии А, к которому, в конечном итоге (при стремящемся к бесконечности времени наблюдения), приходит процесс при данных условиях (температуре и т. д.).
С какой скоростью система приходит к этому равновесному состоянию? Для ответа решим соответствующее дифференциальное уравнение:
dnA/dt = - kА®В nA + kB®A nB | (17.5) |
(второе уравнение, для dnB/dt, писать не надо, т. к. nA+ nB º n0, где n0 — полное число молекул, т. е. dnВ/dt º -dnА/dt). Ответ — позвольте мне написать его без выкладок, а вам я рекомендую сделать их самостоятельно — таков:
nA(t) = [nA(0) — nА¥] exp[-(kА®В + kB®A)t] + nА¥ . | (17.6) |
Здесь nA(t) — число молекул в состоянии А в момент времени t от начала процесса, а nА¥ = n0.[kВ®А / (kА®В + kВ®А)] — конечное, равновесное число молекул в состоянии А.
Значит, видимая (apparent) скорость приближения к равновесию есть
kapp = kА®В + kВ®А, — | (17.7) |
она равна сумме скоростей прямой и обратной реакций. Обратите внимание, что эта скорость зависит только от условий, в которых протекает релаксация, и не зависит от исходного распределения белковых молекул между нативным и денатурированным состояниями [т. е. от исходной доли nA(0)].
В этой суммарной скорости kapp доминирует более быстрая реакция.
Если условия (температура и т. д.) отвечают большей устойчивости нативного состояния, — то в kapp = ku®N + kN®u доминирует ku®N, скорость сворачивания. Если более устойчиво денатурированное состояние, — то в ku®N + kN®u доминирует kN®u, скорость разворачивания.
Большим преимуществом измерения скорости прихода к равновесию, kapp, является то, что она всегда может быть измерена (в отличие от ku®N и kN®u в отдельности), — как тогда, когда более стабильно нативное состояние, так и тогда, когда более стабильно денатурированное.
При измерении kapp строятся так называемые "шевронные графики" (Рис.17-8). Они так называются из-за своей характерной V-образной формы, напоминающей шевроны на военных кителях. Один взгляд на этот график показывает, что скорости сворачивания (ku®N) и разворачивания (kN®u) зависят от концентрации денатуранта противоположным образом. С аналогичной ситуацией мы уже сталкивались, когда изучали температурную зависимость скоростей сворачивания и разворачивания белка. Как и тогда, наблюдаемая здесь противоположность наклонов означает "промежуточность" свойств переходного состояния, их среднее положение между свойствами нативного и развернутого белка.
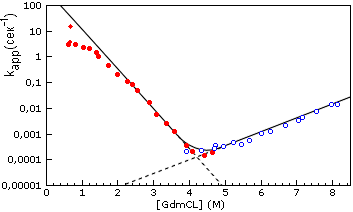
Рис.17-8. "Шевронный график" зависимости kapp = ku®N + kN®u, видимой характерной скорости приближения к равновесию между нативной и развернутой формами белка (лизоцима куриного яйца), от концентрации гуанидингидрохлорида. Заполненные кружки получены при разбавлении раствора GdmCl, в котором изначально находился денатурированный белок, т. е. при полной или частичной ренатурации белка. При этом kapp » ku®N. Пустые кружки получены при добавлении GdmCl к изначально нативному белку, т. е. при полной или частичной его денатурации. При этом kapp » kN®u. Пунктир показывает экстраполяцию величин ku®N и kN®u в области излома шеврона. Отклоняющиеся от экстраполяционной прямой точки в верхней левой части графика (т. е. вдали от точки излома, от точки денатурации белка) свидетельствуют либо о какой-то перестройке переходного состояния, либо о появлении каких-то дополнительных метастабильных интермедиатов (возможно, типа расплавленных глобул), которые могут лежать как на, так и вне основного пути сворачивания. Обратите внимание, что все эти перестройки и/или интермедиаты не повышают скорость сворачивания по сравнению с той, что можно было бы ожидать при неизменности переходного состояния: отклоняющиеся точки лежат ниже интерполяционной прямой. Картинка, с небольшими изменениями (добавлены экстраполяционные пунктиры) взята из T. Kiefhaber Proc. Natl. Acad. Sci. USA (1995) 92:9029-9033.
Действительно, то, что денатурант вообще разворачивает белок, показывает, что он сильнее притягивается к развернутому белку, чем к нативному. То, что денатурант замедляет сворачивание исходно развернутого белка (см. спад скорости в левой части Рис.17-8) показывает, что он сильнее притягивается к исходному, развернутому белку, чем к переходному состоянию; а то, что денатурант ускоряет разворачивание исходно нативного белка (см. подъем скорости в правой части Рис.17-8), — показывает, что он сильнее притягивается к переходному состоянию, чем к исходному, нативному. Это означает, что контакт переходного состояния с денатурантом больше, чем у нативного белка, но меньше, чем у денатурированного. Иначе говоря, приведенный на Рис.17-8 шевронный график показывает, что переходное состояние по степени контакта с растворителем, т. е. по своей компактности находится где-то на полпути между развернутым и нативным состояниями белка. Этот график дает даже больше: так как наклон для kN®u несколько меньше, чем для ku®N, то компактность переходного состояния несколько ближе (в данном случае) к оной у нативного белка, чем у денатурированного.
Рисунок 17-8 относится к лизоциму — белку, денатурация которого гуанидингидрохлоридом приводит прямо к образованию клубка, а не расплавленной глобулы. При умеренных концентрациях денатуранта (при не очень сильном его разбавлении водой) лизоцим сворачивается довольно медленно, за многие минуты. Однако при самоорганизации в почти чистой воде (происходящей при сильном разведении водой крепкого раствора гуанидингидрохлорида, где находились клубкообразные белковые цепи), скорость сворачивания лизоцима максимальна, — и уже слабо зависит от остаточной концентрации денатуранта. При этом в сворачивании лизоцима появляется компактный метастабильный интермедиат типа расплавленной глобулы, — интермедиат, который не наблюдается при повышенных концентрациях денатуранта. Появление компактных метастабильных интермедиатов сворачивания, вообще говоря, всегда "выполаживает" зависимость скорости сворачивания от содержания денатуранта — см. левый край Рис.17-8. Известно, что ренатурация белка, стартующая не от клубка, а от расплавленной глобулы (например, ренатурация карбоксиангидразы), также демонстрирует довольно слабую зависимость скорости ренатурации от концентрации денатуранта. Последнее означает, что переходное состояние в таком сворачивании мало отличается от исходной расплавленной глобулы по компактности.
Иначе говоря, переходное состояние — по "промежуточности" некоторых своих свойств между свойствами нативного и развернутого белка — напоминает расплавленную глобулу. Но это не означает, что переходное — подчеркнем, нестабильное — состояние действительно похоже на расплавленную глобулу. Эксперимент показывает скорее, что переходное состояние можно представить себе (если речь идет о переходе "клубок ® нативный белок") как частично свернутый нативный белок, остальная часть которого все еще находится в денатурированном, клубкообразном состоянии. Вид переходного состояния для переходов типа "расплавленная глобула ® нативный белок" (или "клубок ® расплавленная глобула ® нативный белок") еще не установлен экспериментально, но весьма правдоподобно, что оно включает часть нативной глобулы, в то время как прочая цепь находится в состоянии расплавленной глобулы.
Природу переходного состояния удалось выяснить с помощью белковой инженерии. Применяя множество мутаций, и анализируя соответствующие им изменения в шевронных графиках (см. Рис.17-8, 17-9), удается — ценой колоссального труда — узнать, какие именно остатки вовлечены в "нативоподобную" часть переходного состояния, а какие — нет. Этот метод был разработан А. Ферштом а Англии. Строго говоря, пока что он применяется только к белкам, денатурация которых приводит прямо к образованию клубка, а не расплавленной глобулы.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
