
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
ВВЕДЕНИЕ В ФИЗИКУ БЕЛКА
КУРС ЛЕКЦИЙ
1999-2000 гг.
Алексей Витальевич Финкельштейн
Посвящаю светлой памяти моего деда, биолога,
профессора , комиссара
тифозных бараков 20-го года, несломленного
"вейсманиста-морганиста" 48-го.
ПРЕДИСЛОВИЕ
Предлагаемые вашему вниманию лекции посвящены физике белка, т. е. самым общим проблемам структуры, самоорганизации и функционирования белковых молекул.
Поскольку то, что Вы видите перед собой — запись курса лекций, а не монография и не учебник, здесь неизбежны повторы (и, в частности, — повторы рисунков). Ведь в реальных лекциях невозможны отсылки типа "смотри рисунок 2 и формулу 3 в пред-предыдущей лекции"; однако я приложил все старания к тому, чтобы такие повторы были минимальны.
По ходу курса я не упущу возможности изложить те физические идеи и, в частности, те элементы статистической физики и квантовой механики, которые, с моей точки зрения, абсолютно необходимы для понимания строения и функционирования белков, — и которые "нормальный" студент-биолог либо прочно забыл к четвертому курсу, либо не знал никогда.
А вот из моря информации о функциях белков я зачерпну только то, что абсолютно необходимо для иллюстрации роли, играемой пространственными структурами белков в их биологической — точнее, биохимической — деятельности.
Эти лекции, безусловно, отражают мои личные вкусы и пристрастия.
В них много теории и физических проблем — и лишь необходимый минимум экспериментальных фактов (и почти нет экспериментальной техники). Поэтому они никак не заменяют обычные биофизические и биохимические "белковые" курсы и не должны использоваться как справочник. Говоря о конкретных белках, я даю лишь важнейшие (надо ли говорить — с моей точки зрения!) примеры; таблицы включают лишь абсолютно необходимые данные; рисунки — намеренно схематичны; все цифры — округлены, и т. д.
Кроме того, я не упущу возможности отметить то, что сделали по физике белка мы — в Институте белка РАН. Это, конечно, усилит "личный элемент" лекций — и, я думаю, немного оживит их...
При подготовке компьютерного издания лекций (декабрь 1999 — август 2000) их текст был заново отредактирован.
Лекция 1
Белки — и молекулярные машины, и строительные блоки, и оружие живой клетки. Важнейшая и почти монопольная функция белков — ферментативный катализ химических превращений в клетке и вокруг нее. Кроме того, белки-регуляторы контролируют экспрессию генов, а рецепторные белки (они сидят в липидной мембране) обеспечивают восприятие межклеточных сигналов, часто передаваемых гормонами (тоже белками). Иммунные белки и сходные с ними белки гистосовместимости опознают и связывают "чужие" молекулы, а также "свои" клетки — что и позволяет им (клеткам) правильно размещаться в организме. Структурные белки образуют микрофиламенты, микротрубочки, — а также фибриллы, волосы, шелк и другие защитные покровы; они армируют мембраны и поддерживают структуру клеток и тканей. Транспортные белки переносят (а запасные — запасают) другие молекулы. Белки, переносящие протоны и электроны через мембрану, обеспечивают всю биоэнергетику — поглощение света, дыхание, выработку АТФ. Другие белки, "сжигая" АТФ, обеспечивают механохимическую деятельность, — они работают в мышцах или перемещают элементы клетки.
При всем разнообразии, работа белков всегда базируется на высоко специфическом — как у ключа с замком (точнее: как у гибкого ключа с гибким замком) — взаимодействии белка с обрабатываемой им молекулой. Для специфического взаимодействия необходима достаточно "твердая" пространственная структура. Поэтому биологическая функция белков (как и других важнейших для жизни макромолекул — ДНК и РНК) тесно связана с определенностью их трехмерных структур. Не только разрушение — даже небольшие изменения этих структур часто ведут к утере или резкому изменению активности белков.
Знание молекулярной трехмерной структуры белка необходимо для понимания функционирования белковой молекулы. Поэтому в этих лекциях я буду говорить о физике функционирования белков лишь в конце, сосредоточившись вначале на их структуре, на природе ее стабильности и способности к самоорганизации.
В основе белка лежит полимер — состоящая из аминокислотных остатков полипептидная цепь; это было установлено Э. Фишером в начале века. В начале 50-х годов Сэнгер показал, что белковая цепь имеет уникальную последовательность звеньев — аминокислотных остатков ("остаток" — это то, что осталось от свободной аминокислоты после ее встраивания в белковую цепь). Эта цепь имеет химически регулярный остов ("главную цепь"), от которого отходят разнообразные боковые группы аминокислот — радикалы R1, R2, ..., RM
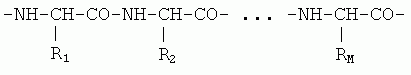
Есть двадцать основных аминокислотных остатков. Их положение в белковой цепи кодируется генами. Однако последующая модификация белка иногда увеличивает разнообразие аминокислот. Кроме того, в некоторые белки включаются разные кофакторы — малые молекулы, ионы, сахара, нуклеотиды, фрагменты нуклеиновых кислот и т. д. В ряде случаев они ковалентно пришиваются к определенным точкам цепи белка, но часто и просто специфически "прилипают" к белку.
В "работающем" белке его цепь свернута строго определенным образом. В конце 50-х Перутц и Кендрью расшифровали пространственное строение первых белков и показали высокую сложность и уникальность их строения. Впервые же (как потом выяснилось!) строгую определенность строения белковых молекул фактически доказал еще в 60-х годах прошлого века Хоппе-Зейлер, получив кристаллы гемоглобина: ведь в кристаллах каждый атом каждой молекулы знает свое место.
Вопрос о том, одинакова ли структура белка в кристалле и в растворе, обсуждался долгие годы, пока ЯМР не показал, что обычно она практически (с точностью до флуктуаций) одна и та же.
Белки "живут" в разном окружении, и их структура несет на себе явную печать этого окружения. Чем меньше воды вокруг белка, — тем невосполнимее разрыв мощных водородных связей, стягивающих его цепь, тем ценнее для белка эти связи (а именно они крепят структуру остова белковой молекулы), тем регулярнее вынуждена быть стабильная структура белка.
Грубо говоря, по "жизненным условиям" и общему типу строения белки можно разбить на три класса.
1) Фибриллярные белки образуют огромные агрегаты; их структура высоко регулярна и держится в основном взаимодействиями между разными цепями.
2) Мембранные белки "живут" в мембране, где нет воды, но части их выступают из мембраны в воду. Внутримембранные части таких белков — как и фибриллярные белки — высоко регулярны и прошиты водородными связями, но размер этих регулярных частей ограничен толщиной мембраны.
3) Водорастворимые, живущие в воде глобулярные белки наименее регулярны (особенно небольшие); их структура держится взаимодействиями белковой цепи с самой собой, причем особенно важны взаимодействия далеких по цепи, но сблизившихся в пространстве углеводородных (гидрофобных — "боящихся воды") групп, а также взаимодействиями белковой цепи с кофакторами.
Наконец, небольшие или бедные углеводородными группами полипептиды могут сами по себе не иметь фиксированной структуры, но приобретать ее при взаимодействии с другими макромолекулами.
Конечно, намеченное выше деление очень грубо. Порой белок может состоять из фибриллярного "хвоста" и глобулярной "головки" (так устроен, например, миозин), и так далее.
Уже известны сотни тысяч белковых аминокислотных последовательностей (для их хранения созданы специальные компьютерные банки — например, SwissProt) и тысячи пространственных структур белков (они хранятся в компьютерном Банке Белковых Структур). Подавляющая часть того, что мы знаем о трехмерных белковых структурах, относится к водорастворимым глобулярным белкам. Для мембранных же и фибриллярных белков расшифрованы лишь считанные пространственные структуры или отдельные фрагменты. Причина проста: водорастворимые белки легче выделять в виде отдельных молекул и их структуру легче изучать и рентгеном — в кристаллах, и спектроскопией ЯМР (ядерного магнитного резонанса) — в растворах. Поэтому, говоря о "структуре белка", "формировании структуры белка" и т. д. — часто, на самом деле, имеют в виду закономерности, доказанные лишь для водорастворимых глобулярных белков. Это надо иметь в виду, читая книги и статьи.
Нековалентные взаимодействия, поддерживающие пространственное строение белка, значительно слабее химических связей, фиксирующих последовательность мономеров — аминокислот — в белковой цепи. Эта последовательность — она называется "первичной структурой белка" (Рис.1-1) — создается в ходе матричного биохимического синтеза согласно "инструкции", записанной в гене.
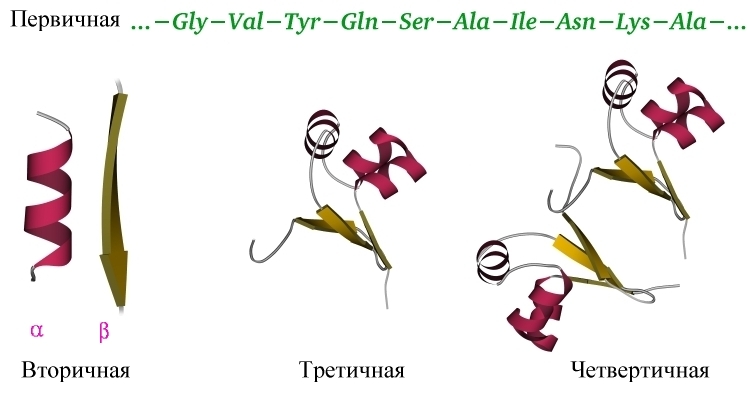
Рис.1-1. Уровни организации белковой структуры: первичная структура (аминокислотная последовательность), вторичная структура (a-спираль и один тяж b-структуры), третичная структура глобулы, сложенной одной цепью, и четвертичная структура олигомерного (в данном случае - димерного) белка.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
