
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
По мере роста информации о пространственном строении белковых молекул становилось все яснее, что существуют какие-то "типовые проекты" строения белковых глобул. Архитектуры вновь расшифрованных белков (или, по крайней мере, их доменов) все чаще и чаще оказывались сходными с архитектурами белков уже известных — но при этом совсем других и по функции, и по аминокислотной последовательности. Поэтому причина сходства структур, видимо, заключается не только в эволюционной дивергенции и не (или не только) в функциональной конвергенции белков, а в ограничении набора укладок какими-то физическими закономерностями.
В 70-х годах стало ясно, что между двумя "традиционными" структурными уровнями (вторичная структура белка и его детальная атомная трехмерная структура) находится промежуточный уровень — "мотив укладки" белковой цепи, определяемый взаимным расположением a - и/или b-участков в глобуле — и что именно на этом уровне проявляется сходство белков, не связанных ни эволюционно, ни функционально. В отличие от детальной, атомной трехмерной структуры, "мотивы укладки" удивительно просты и даже красивы (Рис.15-1).
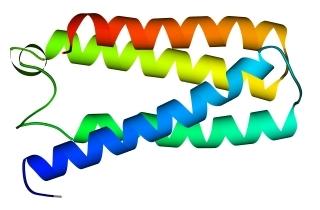


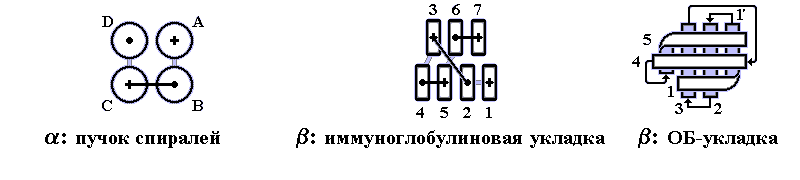
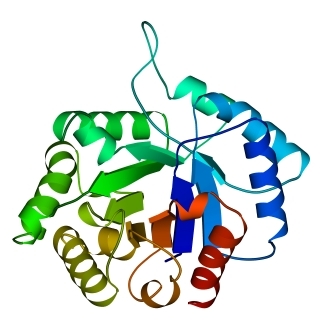
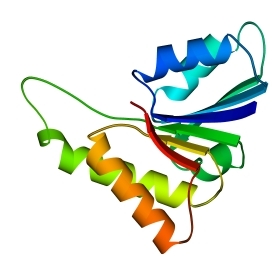
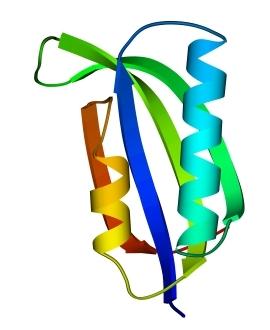
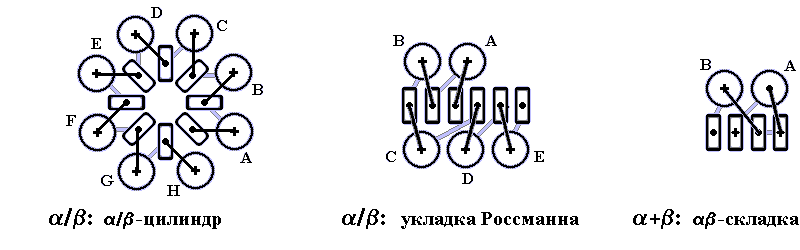
Рис.15-1. Характерные мотивы укладки белковой цепи в a, b, a/b и a+b белках. Внизу — их упрощенные схемы, вид с торца укладки. Обратите внимание на слоевую упаковку a - и b-структур и на то, что каждый слой сложен либо только из a-спиралей, либо только из b-тяжей, но не из a-спиралей и b-тяжей одновременно.
В поисках ответа на вопросы: (1) В чем физическая причина простоты и регулярности типичных мотивов укладки белковой цепи? и (2) Почему одни и те же мотивы встречаются в самых разных белках, и чем замечательны именно эти мотивы? — мы исследуем прежде всего стабильность различных структур. Такой подход оправдывается тем, что одни и те же пространственные структуры белков могут быть получены в результате кинетически совсем разных процессов. Они получаются и in vivo (как в процессе биосинтеза белка на рибосоме, так в процессе транслокации — в более или менее развернутом виде — через мембрану), и in vitro, при сворачивании (ренатурации) целой белковой цепи из развернутого состояния. Это значит, что детальная последовательность действий не играет решающей роли при сворачивании белка.
Начнем с простого вопроса — почему существует слоевое строение глобулярных белков, о котором мы говорили на прошлой лекции. Иными словами, — посмотрим, почему стабильность плотной глобулы требует, чтобы каркас белковой молекулы выглядел бы как компактная упаковка a - и b-слоев, чтобы a - и b-участки шли от одного края глобулы до другого, и чтобы нерегулярные участки не лежали внутри глобулы.
В общем, мы об этом уже говорили. Тут все дело в водородных связях, которые стоят дорого, а потому все должны быть насыщены в стабильной структуре. Доноры и акцепторы таких связей есть в пептидной группе каждого аминокислотного остатка. Насытиться они могут или водой, или при образовании вторичной структуры. Поэтому только вторичные структуры могут не контактировать с водой — лежать внутри глобулы — а содержащие свободные полярные пептидные группы элементы — петли, края b-листов и концы a-спиралей — должны быть на поверхности.
Вытянутые a - и b-структуры должны, ради стабильности глобулы, со всех сторон плотно окружать гидрофобное ядро, создаваемое боковыми группами этих участков, и тем самым отделять его от воды. В то же время a-спирали и b-листы не могут смешиваться в одном слое, — пропадут водородные связи края b-листа. Значит, стабильность глобулы требует образования a-слоев и — отдельно — b-слоев (Рис.15-1). Такие слои (обычно не плоские — скрученные, иногда цилиндрические, а в a-спиральных глобулах — даже квазисферические) действительно, как мы видели, типичны для белковых глобул.
Подавляющее большинство доменов может быть представлено в виде двух-, трех - или (редко) четырехслойных структур, хотя отдельные белки (особенно те, которые содержат металлоорганические комплексы или много S-S связей боковых групп) могут и не вполне удовлетворять этой схеме. Более чем четырехслойных доменов нет — и в принципе ясно, почему. У них внутри, в отдалении от воды находилось бы слишком много остатков, и при типичном для белковых цепей — точнее, для цепей водорастворимых глобулярных белков — соотношении 1:1 между неполярными и полярными остатками многие полярные остатки увлекались бы внутрь белка, что энергетически крайне невыгодно: такой белок не был бы стабильным. Поэтому очень большие единые глобулы "обычного" аминокислотного состава должны быть нестабильны, и большие белки должны разбиваться на субглобулы, домены.
В принципе, можно, видимо, придумать такие аминокислотные последовательности, боковые группы которых как бы "залечат" все разрывы водородных связей между главной цепью и водой, — разрывы, которые последуют за погружением в глобулу края b-листа или петли, так и насытят водородными связями увлеченные внутрь белка полярные боковые группы. Или — придумать последовательности, которые с лихвой заплатят за эти разрывы мощными связями — например, ковалентными (S-S) или координационными (как в металлоорганике). Придумать можно. Но это будут очень специальные, — а значит, очень редкие последовательности...
Может быть, тут-то собака и зарыта, — может быть, "нормальные" глобулярные белки создаются "нормальными" (т. е. сравнительно слабо отобранными), а не "очень редкими" (т. е. сильно отобранными), последовательностями!?
Попробуем взглянуть на первичные структуры белков (Рис.15-2). Статистический анализ показывает, что аминокислотные последовательности водорастворимых глобулярных белков — а о них-то сейчас и идет у нас речь — выглядят как "случайные". То есть в них разные аминокислотные остатки перемешаны примерно так, как можно было бы ожидать при случайной сополимеризации. Конечно, каждая последовательность не есть результат случайного биосинтеза; каждая белковая цепь кодируется геном. Однако аминокислотные последовательности водорастворимых глобулярных белков выглядят как "случайные", — в том смысле, что в них нет ни блочности, характерной для мембранных белков (где явно гидрофобные куски перемежаются с явно гидрофильными), ни периодичности, характерной для белков фибриллярных.
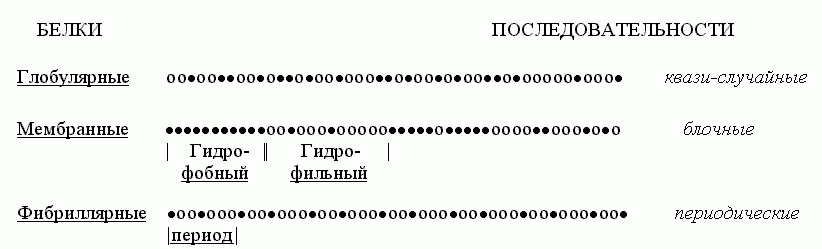
Рис.15-2. Характерные мотивы чередования гидрофобных (·) и полярных (о) аминокислот в первичных структурах водорастворимых глобулярных белков, мембранных белков и фибриллярных белков.
А что такое "выглядеть как случайная последовательность"? Это значит — выглядеть как большинство из всех возможных последовательностей... Значит, рассматривая водорастворимые глобулярные белки, вполне осмысленно ставить вопрос о том, какие стабильные пространственные структуры обычно кодируются самыми массовыми, случайными или похожими на них ("квазислучайными") последовательностями.
Следуя логике такого анализа, один результат мы уже только что получили. Мы выяснили, что типичные упаковки, "штабеля" вторичных структур в глобулярных белках выглядят (Рис.15-1) именно так, как должны выглядеть стабильные упаковки случайных или почти случайных аминокислотных последовательностей.
Пойдем дальше и рассмотрим мотивы укладок белковых цепей.
Как мы уже видели, мотивы укладок белковых цепей часто удивительно красивы. Ход белковых цепей часто напоминает линии, орнаментирующие керамику (Рис.15-3). И, по глубокой мысли Джейн Ричардсон, открывшей это сходство, оно не случайно — так как и линия орнамента, и белковая цепь "решает" одну и ту же задачу — окружить объем (в белке это центр глобулы, ее гидрофобное ядро), избежав самопересечений этой линии.
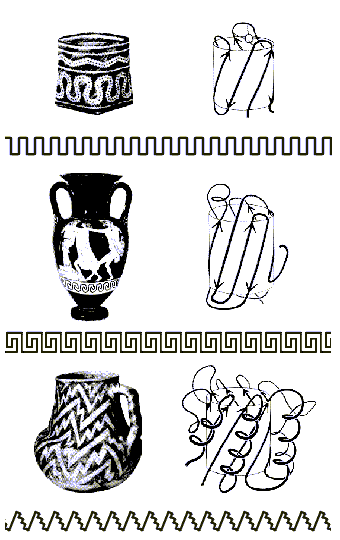
Рис.15-3. Мотивы укладки белковой цепи и орнаменты на индейских и греческих вазах: два решения задачи окружения объема несамопресекающейся линией. Вверху: мотив меандра; в середине: мотив греческого ключа; внизу: мотив зигзага-"молнии". Рисунок взят с обложки Nature, v.268, No.5620, 1977, где была напечатана статья J. Richardson о мотивах укладки белковых цепей.
В белках такой эффект достигается тем, что структурные участки уложены вокруг ядра (или двух ядер; последнее типично для a/b белков), а петли скользят по поверхности ядер — и не перекрывают друг друга (Рис.15-4).
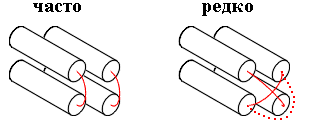
Рис.15-4. Перекрывание петель редко наблюдается в белках, — будь то проход одной петли над другой или обход одной петли вокруг другой.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
