
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Рассмотрим такую задачу формально и попробуем понять, откуда берется наблюдаемая, примерно экспоненциальная зависимость встречаемости разнообразных элементов от их свободной энергии, и что за температура ТС стоит в уравнении (15.1).
Я заранее прошу меня извинить, что все выкладки я дам в самом упрощенном, а значит — не вполне точном виде. Моя цель — дать вам почувствовать, в чем суть дела, не заводя в математические дебри, по которым мы (с и ) в свое время нагулялись вволю...
Итак, пусть De — свободная энергия рассматриваемого элемента в нативном белке, включая его взаимодействия с остальной цепью (например, — свободная энергия лейцина или серина в центре нативной глобулы), отсчитанная от его же свободной энергии в денатурированном белке. Пусть DF — свободная энергия всей остальной цепи в рассматриваемой пространственной структуре, за вычетом свободной энергии в денатурированном белке. Значит, De+DF — разность свободных энергий нативной и денатурированной форм белка. Для того, чтобы нативный белок был стабилен, DF+De должно быть меньше 0, т. е. для стабильного белка
DF < - De. | (15.2) |
Величины DF и De зависят от аминокислотной последовательности полипептидной цепи. Рассмотрим множество последовательностей, оставляющих неизменной величину De [в данном случае — все последовательности с инвариантным Leu (или Ser) в данном месте цепи; при этом, если это место лежит в ядре нативного белка, De = 2 ккал/моль для Leu и около 0 для Ser, в то время как для поверхностных остатков De близко к 0, если денатурированный белок развернут]. Величина же DF будет меняться от последовательности к последовательности.
При этом вероятность того, что DF < - De, есть
| (15.3) |
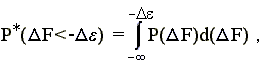
где P(DF) — вероятность появления заданной величины DF в наудачу выбранной случайной последовательности.
Величина DF складывается из свободных энергий множества разных взаимодействий и из энтропий фиксации множества разных аминокислотных остатков. Так как складываемых "случайных" величин много, — то, по "центральной предельной теореме" математической статистики, вероятность (P) того, что DF равна той или иной величине, имеет простую, так называемую Гауссову (см. Рис.15-7) форму:
P(DF) = (2ps2)-1/2 x exp[-(<DF> — DF)2/2s2] . | (15.4) |
Здесь <DF> — средняя (усредненная по всем последовательностям) величина DF, а s — средняя (точнее — среднеквадратичная, усредненная по тем же последовательностям) величина отклонения DF от среднего <DF>.
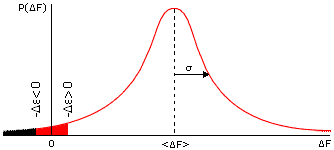
Рис.15-7. Типичный (Гауссов) вид распределения величины свободной энергии DF по различным случайным аминокислотным последовательностям. В DF входит вся свободная энергия укладки белковой цепи (отсчитанная от свободной энергии денатурированного белка), за исключением фиксированной свободной энергии De интересующего нас элемента структуры. Значения DF < - De (т. е. удовлетворяющие условию DF+De < 0) отвечают стабильной укладке. Зачерненная область соответствует значениям DF < - De при De > 0, "красная+зачерненная" облась — при De < 0. Вторая область больше, т. е. больше случайных последовательностей стабилизует укладку цепи, если свободная энергия интересующего нас элемента De < 0, чем если De > 0.
Напомню, что "центральная предельная теорема" описывает ожидаемое распределение суммы большого числа случайных величин, и отвечает на вопрос "какова вероятность того, что эта сумма будет иметь то или иное значение?". У нас "сумма большого числа случайных величин" — это DF (она включает множество взаимодействий в наудачу выбранной, а значит — "случайной" последовательности). Так вот, математика говорит, что для большинства последовательностей величина такой суммы DF должна лежать между <DF> - s и <DF> + s, и что вероятность величины DF круто, экспоненциально падает при удалении DF от <DF> (см. Рис.15-7).
Так как рассматриваемая укладка обычно стабильна при DF < 0 (точнее: она стабильна при DF+De<0, но одно-единственное взаимодействие, создающее De, в данном случае не в счет на фоне множества взаимодействий, создающих DF), и так как подавляющее большинство случайных последовательностей явно неспособно стабилизировать именно рассматриваемую укладку цепи (с чего вдруг, — именно ее из огромного числа возможных укладок?), — то величина <DF> должна быть не только положительной, но и большой — существенно больше, чем s (иначе — был бы велик, порядка 1/2, шанс, что случайная последовательность будет стабилизировать именно рассматриваемую укладку).
Величина (<DF> — DF)2 = <DF>2 — 2<DF>DF — DF2, — то есть при малых (относительно большого <DF>) значениях DF, мы имеем (<DF> — DF)2 » <DF>2 — 2<DF>DF — и, значит,
P(DF) » {(2ps2)-1/2 x exp[ - <DF>2/2s2]} x exp[DF x (<DF>/s2)]. | (15.5) |
При этом [прошу проверить — или (хуже!) поверить] — вероятность того, что DF < - De, есть
| (15.6) |
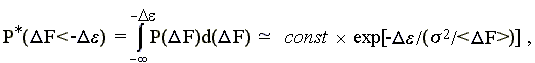
так как не зависящий от переменной DF множитель {(2ps2)-1/2 x exp[-<DF>2/2s2]} выносится за знак интеграла, а интеграл величины exp[DF x (<DF>/s2)] от "минус бесконечности" до "-De" есть (s2/<DF>) ´ exp[-De ´ (<DF>/s2)].
Константа const = {(2ps2)-1/2 xexp[-<DF>2/2s2]} x (s2/<DF>) нас не интересует, а интересующий нас член exp[-De/(s2/<DF>)] демонстрирует, что свободная энергия (De) какого-то элемента белковой структуры экспоненциально изменяет вероятность того, что наугад выбранная аминокислотная последовательность будет стабилизировать данную укладку цепи. Таким образом, увеличение De экспоненциально уменьшает число аминокислотных последовательностей, способных придать стабильность нативной структуре, содержащей элемент с энергией De.
Полученная зависимость имеет экспоненциальную форму (и тем она похожа на формулу Больцмана) — но, в отличие от формулы Больцмана, De здесь делится не на температуру среды (точнее, не на kT), а на непонятную пока величину s2/<DF>.
Что же это за величина? Прежде всего, — отметим, что s2/<DF> не зависит от размера белка. В самом деле, по законам математической статистики, средняя величина (<DF>) пропорциональна числу суммируемых в DF членов (которое, в нашем случае, примерно пропорционально размеру белка), а среднеквадратичное отклонение от среднего (s) пропорционально квадратному корню из числа этих членов, — т. е. De2 тоже примерно пропорционально размеру белка.
То, что s2/<DF не растет с размером белка — очень важно: Это значит, что величину "дефекта" De надо сравнивать — в формуле 15.6 — не с полной энергией белка (которая пропорциональна его размеру), а с какой-то характерной энергией s2/<DF>, т. е. с чем-то вроде средней энергии цепи в расчете на один ее аминокислотный остаток (которая тоже не зависит от размера белка!).
Это, вместе с экспоненциальным видом формулы 15.6, сразу отвечает на вопрос о том, почему De ценой всего в несколько ккал/моль может существенно влиять на встречаемость белковых структур (например — почему лейцина внутри белка на порядок больше, чем серина). Это происходит потому, что число последовательностей, стабилизующих нативную структуру белка, падает в "е" раз (примерно втрое), когда De растет на s2/<DF>.
Итак, величина s2/<DF> — это что-то вроде энергии цепи в расчете на одно ее звено или на одну ее степень свободы. Или (напомню, что kT — это характерная тепловая энергия в расчете на степень свободы) — можно полагать, что s2/<DF> = kTC, где TC — какая-то температура. Какая? В белке есть только одна характерная температура — температура его плавления, денатурации. И мы все время говорили об устойчивости белка к денатурации... Поэтому можно полагать, что величина TC определяется температурой денатурации белка. Иными словами, можно предполагать, что s2/<DF> составляет примерно 0.5 — 1 ккал/моль.
Это можно не только предполагать, но и показать. За словами "можно показать" стоит довольно сложное теорфизическое доказательство — теорема Шахновича-Гутина — от которого я вас избавлю.
Итак, мы разобрались с вопросом о том, почему дефект ценой всего в несколько ккал/моль — на фоне гораздо большей полной энергии белка — может практически запрещать многие мотивы белковых архитектур. Потому, что "дефект" ценой в 1 ккал/моль снижает число "годных" для белка последовательностей впятеро, "дефект" ценой в 2 ккал/моль — раз в двадцать, и т. д.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
