
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Архитектуры белков, особенно глобулярных водорастворимых белков, сложны и разнообразны — в противоположность универсальности двойной спирали ДНК (одноцепочечные РНК здесь как бы занимают промежуточное положение). Однако и в белках прослеживается набор "стандартных" структур, о которых мы будем говорить весьма подробно.
Прежде всего здесь речь идет о регулярных вторичных структурах белка — об a-спирали и b-структуре; a-спирали часто изображаются спиральными лентами (см. Рис.1-1) или цилиндрами, а вытянутые b-структурные участки (слипаясь, они образуют листы) — стрелками (см. Рис.1-1). Вторичные структуры отличаются регулярной, периодической формой (или, как говорят, "конформацией") главной цепи, — при разнообразии конформаций боковых групп.
Укладка вторичных структур одной полипептидной цепи в глобулу называется третичной структурой (Рис.1-1). Объединение же нескольких белковых цепей в "суперглобулу" называется четвертичной структурой белка (Рис.1-1). Пример (в дополнение к приведенному на Рис.1-1 димеру белка cro репрессора): гемоглобин состоит из двух b и двух a цепей (к a - и b-структурам это никакого отношения не имеет!). Если четвертичная структура белка сложена из одинаковых цепей, то она обычно симметрична (и cro репрессор, и гемоглобин здесь не исключение).
Среди третичных структур есть наиболее типичные, с которыми нам предстоит еще познакомиться. Часто такие типичные структуры охватывают не весь глобулярный белок целиком, а лишь его компактную субглобулу — "домен" (Рис.1-2).

Рис.1-2. Доменная структура большого белка сходна с четвертичной структурой, сложенной из малых белков.
Образование трехмерной структуры белка in vivo происходит при биосинтезе или сразу после него. Замечательно, однако, что оно может происходить не только при биосинтезе: 40 лет назад Анфинсен показал, что оно может идти и при "ренатурации" — сворачивании чем-то развернутой белковой цепи in vitro; причем идти совершенно спонтанно — без помощи других макромолекул.
Это значит, что аминокислотная последовательность сама (при подходящей температуре и рН воды!) определяет пространственную структуру белка — т. е. белок способен к самоорганизации.
Опыты Анфинсена принципиально отделили физический процесс — самоорганизацию пространственной структуры — от биохимического синтеза белковой цепи. Стало ясно, что структура белка детерминирована самой его аминокислотной последовательностью, а не навязывается белку клеточной машинерией. По-видимому, основная задача этой машинерии — оберегать сворачивающийся белок от нежелательных контактов (к коим относятся и контакты между отдаленными, образующими разные домены кусками цепи очень больших белков) — ведь in vivo белок сворачивается в клеточном бульоне, где так много разных молекул, так много возможностей прилипнуть к чему-нибудь! Но в разбавленном растворе in vitro белок — по крайней мере, не очень большой — складывается спонтанно, сам по себе.
Строго говоря, белок способен к спонтанной самоорганизации и ренатурации, только если он не подвергся сильной пост-трансляционной модификации — т. е. если его химическая структура не была сильно нарушена после биосинтеза. Например, инсулин (где половина цепи вырезается уже после того, как он свернулся) не способен к ренатурации.
Пост-трансляционные модификации бывают самые разные. Как правило, химические модификации контролируются специальными ферментами, а не "самоорганизуются" в самом белке. Кроме расщепления белковой цепи (протеолиза: он часто способствует превращению зимогена в активный энзим), наблюдается модификация концов цепи, ацетилирование, гликозилирование, пришивание липидов в определенные точки цепи, фосфорилирование определенных боковых групп, и т. д., и т. п. Недавно был обнаружен даже "сплайсинг" белковых цепей (спонтанное вырезание куска цепи и склейка образовавшихся при этом концов). Изредка наблюдается и спонтанная циклизация кусочков белковой цепи.
Особо надо отметить завязывание S-S связей между серусодержащими остатками цистеина белковой цепи: "правильные" S-S способны (в подходящих условиях in vitro) к спонтанной самоорганизации, хотя in vivo их образование катализируется специальным ферментом, дисульфидизомеразой. Как правило, эти связи завязываются только в секретированных белках (внутри клетки нет кислорода, и потому — неподходящий окислительный потенциал); при этом правильно завязанные S-S связи отнюдь не мешают, а скорее способствуют ренатурации белка.
Итак, аминокислотная последовательность определяет пространственную структуру белка, а структура эта уже определяет его функцию, — т. е. с кем этот белок взаимодействует и что при этом делает. Но, сказав это, надо иметь в виду масштабы. Если обрабатываемая молекула велика — с ней может взаимодействовать весь белок, и тогда вся его форма важна для его функции. Если же обрабатываемая белком молекула мала, то именно мелкомасштабные детали небольшого участка поверхности белка определяют его функцию, а остальное его тело лишь фиксирует эти критически важные детали. Так или иначе, специфичность белка подразумевает его твердость.
Здесь необходимо сделать несколько замечаний.
Первое. Глядя на Рис.1-1, можно подумать, что внутри белка много пустого пространства и, следовательно, что он "мягок". На самом деле это вовсе не так! Белок тверд — белковая цепь упакована плотно, атомы упираются в атомы (см. левую часть Рис.1-3). Но вот только анатомию белка, его скелет, его внутреннее строение неудобно изучать на полной атомной модели — внутреннее устройство белка можно увидеть, лишь сделав атомы "прозрачными" и обратив основное внимание на ход белковой цепи (см. правую часть Рис.1-3 и, в особенности, Рис.1-1, 1-2, где боковые группы "сбриты", а элементы вторичной структуры — выделены). На полноатомной же модели белка (Рис.1-3) даже не видно, что белок образован полимерной цепью, — видна лишь поверхность глобулы, похожая на картофелину. Однако полноатомная модель нужна для изучения функции белка — специфику его работы определяют именно физико-химические и геометрические свойства поверхности глобулы, этой "картофелины", а дело скелета белка — создать и фиксировать эту поверхность.
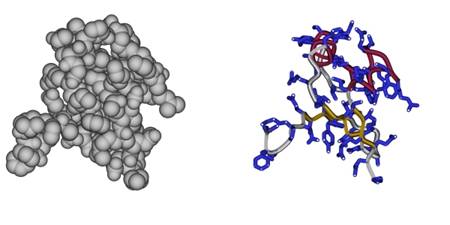
Рис.1-3. Атомная модель белковой глобулы (слева) и ее скелетная модель (справа).
Второе. В состав белка — помимо самой белковой цепи - часто входят кофакторы (Рис.1-4), т. е. относительно небольшие молекулы, принимающие участие в функционировании, а порой и в формировании структуры белковой молекулы. Эти кофакторы могут присоединяться к белку химическими связями, а могут просто прилипать к определенным местам белковой глобулы.
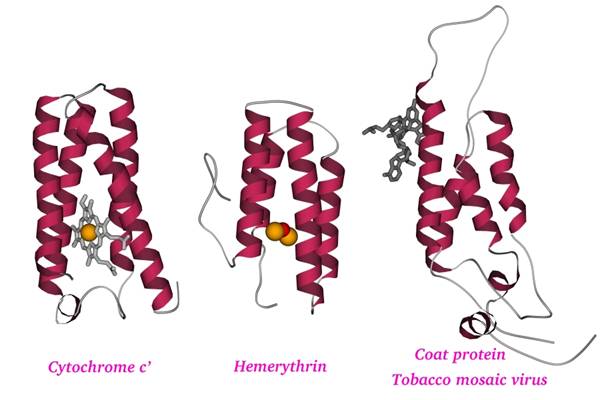
Рис.1-4. Три сходных по форме, но разных по функции a-спиральных белка: цитохром c', миогемэритрин и белок оболочки вируса табачной мозаики. Показана как белковая цепь, так и кофакторы: скелетные модели — гем (в цитохроме) и фрагмент РНК (в белке оболочки вируса), оранжевые шарики — ионы железа (в геме цитохрома и в миогемэритрине), красный шарик — связанный железом кислород (в миогемэритрине).
Третье. Твердый белок ("апериодический кристалл", говоря словами Шредингера) ведет себя именно как кристалл при изменении внешних условий (например, при повышении температуры), — то есть он "терпит" до каких-то пор, а потом разом плавится, — а не теряет своей формы и твердости постепенно, как стекло. Это фундаментальное свойство белков тесно связано с надежностью их работы: как электрическая лампочка, белки ломаются по принципу "все или ничего", а не постепенно (последнее привело бы к ненадежности их действия — к расплыванию специфичности и т. д.).
И последнее. Говоря о твердости, надо различать сравнительно небольшие и действительно твердые однодоменные белки (они состоят из одной компактной глобулы), — и более крупные белки, имеющие либо доменную (Рис.1-2), либо четвертичную (Рис.1-1) структуру: слагающие их субглобулы могут сдвигаться и раздвигаться, а также слегка, как твердое тело, деформироваться (но не полностью перестраиваться!) при функционировании белка.
Обычно сходные по внутренней структуре (по анатомии) белки делают одно и то же. Однако порой одна и та же, в общем, пространственная структура может обеспечивать совсем разные функции. Так, из трех сходных по "грубой" пространственной структуре белков, — один (цитохром) связывает электрон, другой (миогемэритрин) связывает кислород (эти функции немного похожи — обе они включены в цепь окислительных реакций), а третий (белок облочки вируса) связывает гораздо более крупные молекулы — РНК, а также другие оболочечные белки (Рис.1-4).
Верно ли обратное, т. е. определяет ли функция белка его структуру? Здесь тоже прослеживаются некие частные корреляции, но в общем такое определяющее влияние прослеживается только на очень грубом уровне, — уровне, определяемом "средой обитания" белковых функций (так, белки, несущие структурную функцию — например, те, из которых слагаются волосы или фибриллы — как правило, фибриллярны; рецепторы — это мембранные белки, и т. д.). Но часто это совсем неверно на уровне архитектуры белка. Так, две сериновые протеазы (трипсин и субтилизин) имеют одну и ту же каталитическую функцию и даже почти одинаковую специфичность — при совершенно разной (Рис.1-5) внутренней структуре глобул [эти белки сходны не более чем тюлень и жук-плавунец: лишь их (белков) "плавники", — т. е. их активные центры, включающие по полудюжине аминокислотных остатков из пары сотен звеньев цепи, — устроены одинаково, а все остальное — по-разному].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
