
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Итак, чтобы разрешить "парадокс Левинталя" и показать, что самую стабильную структуру белковой цепи можно найти за разумное время, мы можем рассматривать только скорость сворачивания этой структуры цепи вблизи точки ее термодинамического перехода, причем перехода типа "все-или-ничего", в клубок ("клубок" — это сумма всех развернутых конформаций белковой цепи). Иными словами, нам достаточно рассмотреть случай, когда самая стабильная укладка цепи лишь чуть-чуть более стабильна, чем клубок, а все прочие формы белковой цепи термодинамически нестабильны. В окрестности этой точки рассматривать сворачивание белка наиболее просто, т. к. здесь нет стабильных интермедиатов сворачивания. Такие интермедиаты, как о том говорилось на прошлой лекции, появляются лишь тогда, когда нативная структура становится много стабильнее клубка. При этом сворачивание белка достигает максимальной скорости, но его анализ усложняется. Поэтому мы ограничимся окрестностью точки равновесия нативной структуры с клубком, где белок сворачивается пусть не максимально быстро, но максимально просто. Причем мы должны рассматривать только такие аминокислотные последовательности, которые обеспечивают наличие большой "щели" между энергией самой стабильной структуры цепи и энергиями всех прочих ее укладок (Рис.18-1): как вы помните, статистическая физика гетерополимеров показывает, что фазовый, типа "все-или-ничего" распад нативной глобулы требует наличия такой щели.
Рис.18-1. Грубое схематическое изображение энергетического ландшафта белковой цепи: на рисунке мы можем изобразить только две координаты (q1 и q2), описывающих конформацию цепи, тогда как реальная конформация белковой цепи описывается сотнями координат. Широкая щель между глобальным энергетическим минимумом и прочими энергетическими минимумами необходима для того, чтобы стабильная укладка цепи разрушалась бы только путем термодинамического перехода типа "все-или-ничего"; это, в свою очередь, обеспечивает надежность функционирования белка, — по принципу "все-или-ничего", как у электрической лампочки.
Я собираюсь показать, что при таких условиях самая стабильная структура небольшого белка или домена автоматически становится центром "быстрых" путей сворачивания, и потому должна сворачиваться за биологически разумное время — секунды или минуты.
Для того чтобы доказать, что самая стабильная белковая структура должна сворачиваться быстро, достаточно показать, что к ней всегда ведет по крайней мере один "быстрый" путь сворачивания. Наличие не одного, а многих путей сворачивания может только ускорить процесс. Напомню, что, в рассматриваемых нами условиях, — вблизи точки термодинамического перехода типа "все-или-ничего" между самой стабильной структурой цепи и клубком, — никакие другие ("полусвернутые") состояния не могут служить ловушками: они не могут "впитать" сворачивающиеся цепи просто в силу малости своей суммарной стабильности. Полезная аналогия здесь, — просачивание воды через трещины в стенке, разделяющей два бассейна: если "емкость" трещин мала, т. е. они не способны впитать всю воду, то каждая новая трещина в стенке может только ускорить наполнение второго бассейна, — так что, рассмотрев просачивание воды через одну трещину, мы оценим минимальную скорость его наполнения.
Чтобы путь был быстр, каждый шаг на этом пути должен проходиться быстро, таких шагов должно быть не слишком много, и — главное! — этот путь не должен преграждаться "барьером" в виде высокой свободной энергии на одной из стадий сворачивания.
Так как время фиксации одного звена мал![]() (~1 нс., судя по измеренной скорости роста a-спиралей в белковых цепях), — то белок, фиксируя одно свое звено за другим, сворачивался бы мгновенно (100-звенная цепь — за ~100 нс.), если бы при этом он не должен был преодолевать свободно-энергетический барьер.
(~1 нс., судя по измеренной скорости роста a-спиралей в белковых цепях), — то белок, фиксируя одно свое звено за другим, сворачивался бы мгновенно (100-звенная цепь — за ~100 нс.), если бы при этом он не должен был преодолевать свободно-энергетический барьер.
Итак: главный вопрос, на который надо ответить — высок ли барьер на пути, ведущем к самому стабильному состоянию белковой цепи?
Сворачивание белковой цепи ведет к падению ее энтропии (из-за роста упорядоченности цепи) и энергии (из-за образования в цепи контактов между сближающимися звеньями). Падение энтропии повышает, а падение энергии понижает свободную энергию цепи. Если, по ходу сворачивания, цепь должна очень близко подойти к своей финальной структуре перед тем, как начнут возникать стабилизирующие эту структуру контакты (т. е. цепь должна потерять почти всю свою энтропию перед тем, как начнет выигрываться энергия), — то повышение свободной энергии на первом этапе сворачивания будет пропорциональным числу звеньев в цепи, т. е. очень большим, а сворачивание цепи — страшно медленным (как вы помните, — согласно химической кинетике, время протекания процесса экспоненциально зависит от достигаемого по его ходу максимального повышения свободной энергии). Именно такая картина (проигрыш всей энтропии до начала выигрыша энергии) лежит в основе "парадокса Левинталя", утверждающего, что белковая цепь никак не может — даже за время жизни Вселенной — найти свою самую стабильную структуру.
Напротив, если путь сворачивания таков, что по ходу его падение энтропии практически тут же компенсируется падением энергии, — то он не перекрыт высоким свободно-энергетическим барьером, и сворачивание идет быстро. Именно такая картина, как мы убедимся, и имеет место.
При наличии барьера характерное время протекания процесса оценивается, исходя из классической теории переходного состояния, как
ВРЕМЯ ~ t / exp(-F#/RT) . | (18.1) |
Здесь t ~ 1 нс. — характерное время одного шага процесса сворачивания белка, Т — абсолютная температура, R — газовая постоянная, а F# — высота максимума свободной энергии на пути сворачивания (иными словами, свободной энергии перех![]() дного состояния) относительно свободной энергии исходного (клубкового) состояния цепи.
дного состояния) относительно свободной энергии исходного (клубкового) состояния цепи.
Рассмотрим изменение энергии DE, энтропии DS и результирующей свободной энергии DF = DE - TDS по ходу изображенного на Рис.18-2 последовательного сворачивания белка. На каждом шаге этого пути одно звено цепи извлекается из клубка и занимает то положение, в котором оно находится в финальной (самой стабильной) структуре глобулы. Такой процесс может показаться несколько искусственным (откуда звено знает свое место в финальной структуре?). Однако это впечатление исчезает, если заметить, что мы таким образом просто просматриваем "кино" распада стабильной структуры белка в обратном направлении. То есть такой путь сворачивания есть — среди множества других путей сворачивания. И как мы уже договорились, — для минимальной оценки скорости сворачивания, достаточно оценить скорость сворачивания по одному пути.
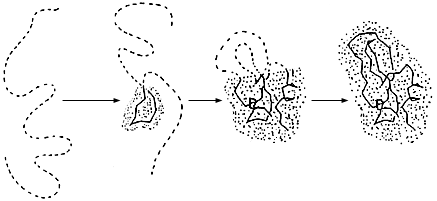
Рис.18-2. Один из возможных путей последовательного сворачивания белка. Пунктиром показана цепь, остающаяся в неупорядоченном состоянии — "клубке". Выделенная точками область соответствует уже обретшей свою финальную конформацию части белковой глобулы. Жирной линией выделена главная цепь уже свернувшейся глобулы, а ее боковые группы не показаны для упрощения рисунка. Все промежуточные состояния имеют высокую свободную энергию и потому не накапливаются при сворачивании и не могут наблюдаться непосредственно.
По мере роста куска финальной глобулы при ее последовательном сворачивании, — в ней одно за другим восстанавливаются взаимодействия, стабилизирующие финальную структуру. Если растущая структура все время остается более или менее компактной (а именно такого типа пути сворачивания, с изображенными на Рис.18-2 компактными интермедиатами, нас и должны интересовать), то число этих взаимодействий будет расти (а их энергия, соответственно, будет падать) почти пропорционально числу n фиксируемых в глобуле звеньев (Рис.18-3а).
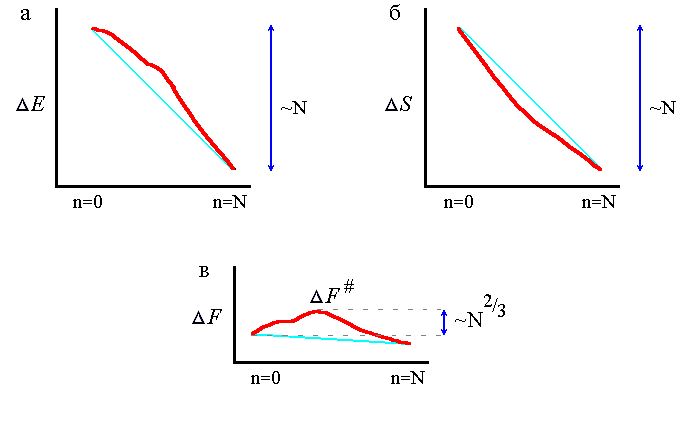
Рис.18-3. Изменение энергии DE (а), энтропии DS (б) и свободной энергии DF=DE-TDS (в) цепи по мере последовательного сворачивания белка (n=0: клубок; n=N: финальная структура) вблизи точки термодинамического равновесия между финальной структурой и клубком. Тонкой линией показана линейная (по n) часть изменения DE(n), DS(n) и DF(n). Полное изменение DE(N) и DS(N) примерно пропорционально N, т. е. числу звеньев в цепи, а максимальное отклонение показанных жирной линией величин DE(n) и DS(n) от линейного (по n) хода относительно мал![]() , — оно пропорционально всего N 2/3. В результате, так DF(N) и, вместе с ней, линейная часть DF(n) = DE(n) - TDS(n) очень мала вблизи точки равновесия, — то DF#, максимальная величина свободной энергии на пути сворачивания, также пропорциональна всего N 2/3.
, — оно пропорционально всего N 2/3. В результате, так DF(N) и, вместе с ней, линейная часть DF(n) = DE(n) - TDS(n) очень мала вблизи точки равновесия, — то DF#, максимальная величина свободной энергии на пути сворачивания, также пропорциональна всего N 2/3.
Правда, в начале сворачивания падение энергии несколько замедлено, так как прилипание звена к поверхности маленькой глобулы дает, в среднем, меньше контактов, чем прилипание к поверхности большой. В результате, возникает нелинейный поверхностный (т. е. пропорциональный n2/3) член в энергии DE растущей глобулы. Таким образом, максимальное отклонение от линейного падения энергии составляет величину порядка N 2/3, где N — число звеньев в белковой цепи. Это отклонение, очевидно, мало по сравнению с полным падением энергии при сворачивании цепи, которое составляет величину порядка N (точнее — порядка Ne, где e —среднее изменение энергии остатка при его переходе из клубка в нативную глобулу).
Во-вторых, по ходу роста глобулы падает энтропия цепи — примерно пропорционально числу фиксированных в глобуле звеньев (Рис.18-3б). Правда, в начале сворачивания энтропия может падать несколько быстрее из-за образования замкнутых петель, свободно торчащих из растущей глобулы (Рис.18-4). В результате, возникает нелинейный (поверхностный) член в энтропии DS этой растущей глобулы, составляющий (так же, как поверхностный член в DE) величину порядка N 2/3, — что существенно меньше, чем само падение энтропии, составляющее (как и падение энергии) величину порядка N.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
