
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
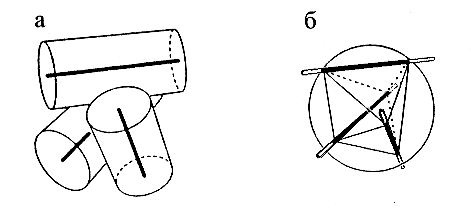
Рис.14-6. Построение многогранника для описания упаковки спиралей. (а) Компактная упаковка трех a-спиралей (цилиндров — 10 ![]() в диаметре — с осями). (б) Построение многогранника: центр упаковки окружается сферой радиусом в 10
в диаметре — с осями). (б) Построение многогранника: центр упаковки окружается сферой радиусом в 10 ![]() ; ее пересечение с осями спирали дает вершины многогранника. На трех из его ребер лежат оси спиралей. Те части ребер, что лежат внутри сферы, оставлены темными. Точки пересечения осей со сферой образуют вершины многогранника. Каждой вершине соответствует одна половинка одной спирали. Оси спиралей образуют часть ребер многогранника, а прочие ребра описывают контакты спиралей.
; ее пересечение с осями спирали дает вершины многогранника. На трех из его ребер лежат оси спиралей. Те части ребер, что лежат внутри сферы, оставлены темными. Точки пересечения осей со сферой образуют вершины многогранника. Каждой вершине соответствует одна половинка одной спирали. Оси спиралей образуют часть ребер многогранника, а прочие ребра описывают контакты спиралей.
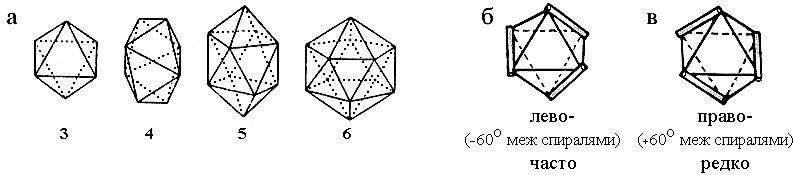
Рис.14-7. Квазисферические многогранники (а), описывающие компактные укладки трех, четырех, пяти и шести спиралей. Большее число спиралей не может уложиться вокруг округлого ядра. Каждый многогранник описывает несколько типов укладок, т. е. типов "штабелей" спиралей, соответствующих различным размещениям осей спиралей на его ребрах. Таких укладок — две для трехспирального комплекса [(б): лево-, и (в): правозакрученный (как на Рис.14-3, 14-4) пучок], десять — для четырехспирального, десять — для пятиспирального, и восемь — для шестиспирального комплекса ("штабеля" для четырех- — шестиспиральных глобул не показаны, но их, при желании, легко построить самостоятельно, разместив спирали — всеми возможными способами — на ребрах многогранника так, чтобы каждая вершина многогранника соответствовала бы одному концу одной спирали). Те упаковки, где межспиральные углы способствуют плотному контакту спиралей — см. Рис.14-9 — встречаются в белках чаще, чем прочие.
Интересно, что в наблюдаемых архитектурах a-спиральных белков вдоль ребер квазисферических многогранников идут не только спирали, но и — как правило — соединяющие их нерегулярные петли (ср. Рис.14-6 и 14-7). Иными словами, в типичном случае белковая цепь как бы обволакивает свое гидрофобное ядро, следуя по непрерывной цепочке ребер квазисферического многогранника.
Обратимся теперь к вопросу о том, как создается плотная упаковка в белковой глобуле. То, что такая упаковка существует, следует из экспериментов, показавших, что белок так же плотен и так же тверд, как органический кристалл. Однако еще предстоит объяснить, как достигается такая упаковка, — слишком уж сложны по форме и разнообразны боковые группы белковой цепи.
Точнее, принцип создания плотной упаковки — и то в самых общих чертах — более или менее ясен только для a-спиралей, почему и уместно рассмотреть этот вопрос именно сейчас.
Первая модель плотной упаковки a-спиралей, упаковка по принципу "выступы (боковые группы) во впадины (между боковыми группами)", была предложена Криком в 1953 г., еще до расшифровки трехмерной структуры первого белка. Затем она была независимо развита Ефимовым и группой Чотиа-Левитт-Ричардсон, и к настоящему времени приобрела вид модели "хребты (боковых групп) в лощины (между таковыми)".
Согласно этой модели, боковые группы на поверхности спирали образуют выступы, создающие разделенные лощинами хребты. "Хребты и лощины" несколько лучше описывают реальность, чем "выступы и впадины", так как разворот одного выступа (одной боковой цепи) в сторону другого (другой боковой цепи) может сделать тот или другой "хребет из выступов" более отчетливым. Хребты (и идущие вдоль них лощины) бывают двух типов. Хребты типа "+4" создаются боковыми группами остатков, расположенных в цепи под номерами "i", "i+4", "i+8", и т. д. (иными словами — с периодом 4), хребты типа "+3" создаются боковыми группами номер "i", "i+3", "i+6", и т. д. (т. е. с периодом 3). Рисунок 14-8 показывает, что эти хребты образуют разного знака углы с осью спирали.
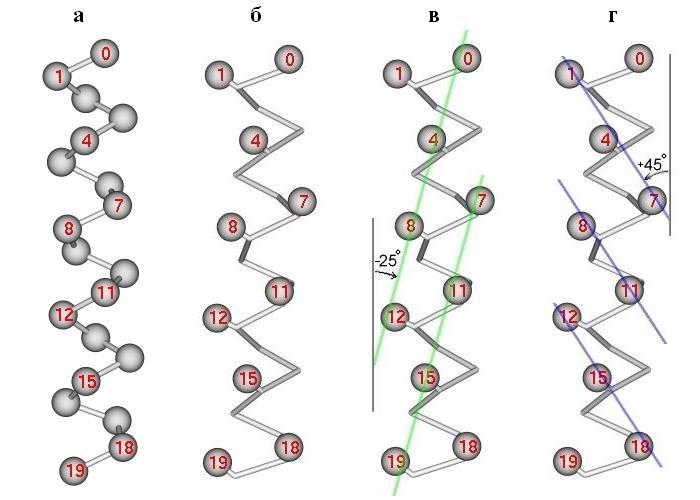
Рис.14-8. a-Спираль; отмечены Сa-атомы (а) и Сb-атомы (б - г). Нумерованные остатки обращены к читателю. Показаны два сорта хребтов (тонкие линии на лицевой поверхности спирали) из сближенных боковых групп (в, г). Хребты из боковых групп "i"-"i+4"-"i+8"... идут под углом -25о к оси спирали (в), хребты из групп "i"-"i+3"-"i+6"... — под углом +45о (г); на рисунке углы представляются меньшими, так как типичные хребты проходят через массивные боковые группы, а на рисунках в, г - через центры Cb-атомов.
При плотной упаковке хребты одной спирали входят в лощины другой. При этом есть две основные возможности:
А) Хребты "+4" одной спирали входят в лощины между такими же хребтами "+4" другой (Рис.14-9а) (здесь для получения плотной упаковки спираль II накладывается на спираль I и поворачивается, пока хребты "+4" обеих спиралей не станут параллельно друг другу). При такой упаковке угол между осями спиралей близок к -500. Этот угол наиболее характерен для контактов спиралей в a-спиральных глобулах. Он также типичен для контакта спиралей в a/b и a+b белках, о которых речь пойдет ниже. Дело в том, что при этом контакте спиралей скрученность слоя a-спиралей (угол скручивания в нем близок к -500/10![]() , где -500 — угол между осями соседних спиралей, а 10
, где -500 — угол между осями соседних спиралей, а 10![]() — ширина a-спирали) неплохо стыкуется с типичной скрученностью b-листа (имеющем тот же угол скручивания, -250/5
— ширина a-спирали) неплохо стыкуется с типичной скрученностью b-листа (имеющем тот же угол скручивания, -250/5![]() , где -250 — угол между осями соседних b-тяжей, а 5
, где -250 — угол между осями соседних b-тяжей, а 5![]() — ширина b-тяжа).
— ширина b-тяжа).
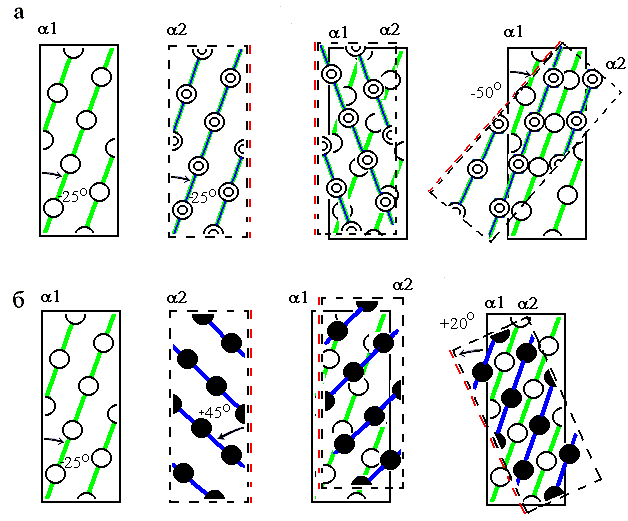
Рис.14-9. Два основных способа плотной упаковки боковых групп при контакте спиралей: под углом -50о (а) и +20о (б). Мы смотрим на зону контакта сквозь одну спираль (сквозь перевернутую вдоль своей оси a2). Остатки "нижней" спирали a1 изображены более светлыми, а верхней (a2) — более темными.
Б) Хребты "+3" одной спирали входят в лощины между хребтами "+4" другой (Рис.14-9б). При такой упаковке угол между осями спиралей близок к +200. Этот угол наиболее характерен для контактов спиралей в пучках — и в a-спиральных глобулах, и в фибриллярных, и в мембранных белках.
Кроме того, хребты "+3" одной спирали могут входить в лощины между такими же хребтами "+3" другой, образуя очень короткий контакт почти перпендикулярных спиралей. В силу малости размера этого контакта, он на рисунке 14-9 не показан, — хотя такой "перпендикулярный" контакт спиралей довольно типичен для a-спиральных глобул.
Заключая описание плотной упаковки, надо заметить, что реальные отклонения от приведенных выше "идеальных" углов весьма велики, так как боковые группы сильно варьируют по размеру. По той же причине в b-структуре (где боковые группы выступают меньше — поверхность b-листа довольно плоская, а спирали — выпуклая) картина типа проникновения хребтов в лощины там совсем смазана и наблюдается лишь в отдельных случаях.
Как согласуется плотная упаковка спиралей с моделью размещения спиралей на ребрах квазисферических многогранников, о которой речь шла выше? Оказывается, довольно любопытным образом. Те "многогранные" упаковки, где углы между спиралями близки к -500 и/или +200, требующимся для плотного контакта спиралей, — эти упаковки встречаются часто; а прочие — редко (но тоже встречаются). Так, из двух изображенных на Рис.14-7 трехспиральных упаковок одна, левозакрученный пучок, приводит к межспиральным углам в -600 (что близко к углу -500, требующемуся для плотной упаковки, см. Рис.14-9а), и такой трехспиральный пучок встречается часто; а другая, правозакрученный пучок, приводит к межспиральным углам в +600 (что далеко от всех углов — -500, +200, 900 — опримальных для плотного контакта), — и такой трехспиральный пучок встречается на порядок реже.
Обратимся теперь к "смешанным" белкам, сложенным из b-листов и a-спиралей. Для них характерна слоистая структура, причем a-спирали и b-участки не могут лежать в одном листе — это привело бы к энергетически невыгодной дегидратации водородных связей на краю b-листа (Рис.14-10).
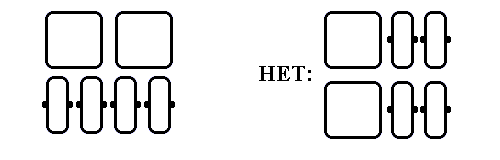
Рис.14-10. Слоистая структура смешанных (a/b, a&b и a+b) белков. Вид с торца b-листа. a-спирали (квадратики - для подчеркивания плотности упаковки глобулы) и b-тяжи (прямоугольники) не могут лежать в одном слое — это привело бы к дегидратации водородных связей (черные точки) на краю b-листа.
Различают a/b ("a дробь b") и a+b ("a плюс b") белки (точнее — домены). Иногда их объединяют в общий разряд a&b ("a и b") белков.
В a/b доменах b-структура параллельна, и a-спирали также параллельны друг другу (и антипараллельны b участкам), а характерное чередование a и b участков в цепи имеет вид - a-b-a-b-a-... .
Есть два характерных мотива строения a/b белков: a/b цилиндр, где b-цилиндр лежит внутри цилиндра, сложенного из a-спиралей (Рис.14-11а), и "укладка Россманна", где более или менее плоский (не считая обычного — правого, если смотреть вдоль b-тяжей — пропеллерного скручивания) b-слой лежит между комплементарно ему скрученными слоями a-спиралей (Рис.14-11б). В отличие от ранее рассмотренных доменов, a/b домены обычно имеют два гидрофобных ядра: в укладке Россманна — между b-листом и каждым из слоев спиралей; в a/b цилиндре — внутри b цилиндра (меньшее ядро) и между b и a цилиндрами (большее).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
