
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Таким образом — "свободная энергия" — это естественное расширение обычного понятия "энергии" на случай, когда система обменивается теплом с окружающей средой. Напомню: если тело не обменивается теплом со средой, его стабильное состояние отвечает минимуму его энергии (или, проще: все, что может упасть — падает). А окружающее тепло делает стабильными определенные флуктуации, прыжки — такие, которые минимизируют свободную энергию тела. То есть приток тепла возбуждает молекулы системы, они начинают захватывать многочисленные состояния с более высокой энергией (т. е. их энтропия растет) — и, в результате, молекулы воздуха летают, а не падают на землю.
Спустимся, однако, с Гималаев к белкам.
Итак — каков же баланс энергий, энтропий и свободных энергий в разобранном нами примере образования водородной связи в белковой цепи? Для полной наглядности, сравним ход процесса в разных условиях:
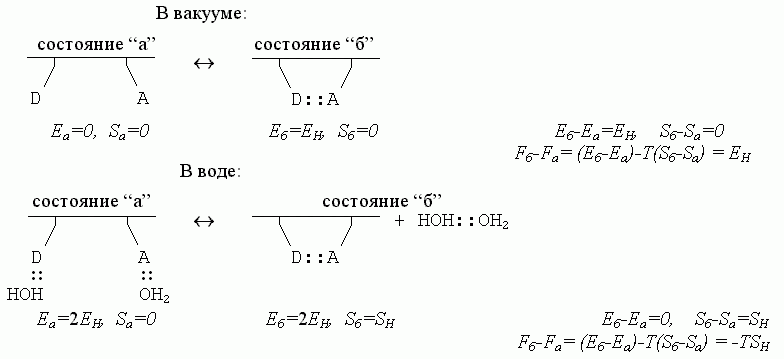
Здесь EH < 0 - энергия Н-связи, а SH > 0 — энтропия перемещений и вращения свободного тела, т. е. свободной молекулы Н2О или свободного димера НОH::OH2. Водородные связи — воды с водой, воды с белком — стабильны, когда EH -TSH < 0 (а если EH -TSH > 0, т. е. если они нестабильны, — то есть в этом случае перед нами была бы уже не жидкая вода, а пар).
Сравнение приведенных выше схем показывает, что водородные связи в белковой цепи как бы становятся — за счет изменения энтропии связывающейся воды — менее стабильными в водном окружении, чем в вакууме; действительно, в водном окружении их свободная энергия есть Fб-Fа = - ТSH , — то есть она меньше, чем в вакууме, где Fб-Fа=EH.
Еще раз хочу подчеркнуть, что причина этого видимого ослабления — в том, что в водном окружении образовавшаяся внутри белковой цепи Н-связь замещает собой связь цепи с водой. И по той же причине водородные связи, стабилизующие структуру белка в воде носят энтропийную, а не энергетическую природу: энергии двух состояний цепи (с внутрицепочечной связью и без нее) примерно равны, и из этих двух состояний с примерно равной энергией стабильнее та, где выше энтропия, т. е. где больше число микросостояний. А их больше у оторванной молекулы воды, чем у связанной.
Обратите внимание: водородные связи в белковой цепи (в водном окружении) носят энтропийную, а не энергетическую природу именно потому, что энергия Н-связей очень высока: раз так, "свободные" от связей в белке доноры и акцепторы водородных связей в цепи всегда на деле не свободны от всех связей вообще, а связаны с водой. Оторвавшиеся же от белка — при образовании внутрибелковой Н-связи — молекулы Н2О тут же связываются друг с другом, так что энергия компенсируется, и свободно-энергетический выигрыш Н-связей в белке идет только от множественности возможных микросостояний оторвавшихся молекул воды. Правда, чтобы связаться друг с другом, молекулы Н2О должны пожертвовать часть своей обретенной свободы, часть энтропии — но лучше уж потерять небольшую энтропию, чем большую энергию!
То, что (1) молекулы воды сильно связаны друг с другом водородными связями; и (2) то, что эти связи образуются только при определенной взаимной ориентации молекул Н2О — определяет специфику воды как растворителя. Это приводит к разным интересным эффектам. Подробнее мы об этом поговорим в двух следующих лекциях.
Лекция 5
В первой части этой лекции я хочу продолжить разговор о термодинамике. Это нам пригодится, когда мы будем потом обсуждать специфику воды как растворителя. Свободная энергия погружения разных молекул в воду играет в этом обсуждении ключевую роль.
Для изучения свободной энергии погружения молекулы в воду берут закупоренную пробирку, наполовину заполненную водой и наполовину — паром, воздухом или органическим растворителем, и смотрят, как изучаемые молекулы распределились между этими двумя фазами.
Мы уже видели, что разность свободных энергий F определяет предпочтительное состояние рассматриваемой системы (в данном случае — предпочтительное местонахождение рассматриваемой частицы) — согласно формуле
[Вероятность быть в б]:[вероятность быть в а] = exp[-(Fб-Fа)/kT] | (5.1) |
Свободная энергия F = E — TS складывается из энергии системы E и ее энтропии S. С энергией все, видимо, понятно. С температурой — понятно скорее интуитивно, и к ней мы еще вернемся. С энтропией — сложнее, и хочется повторить еще раз.
В простейшем случае частицы в сосуде, который мы, если вы помните, рассмотрели, S = k lnV, где V — доступный для частицы объем.
В чем смысл вхождения в свободную энергию этой энтропии (в виде члена -TS)? В том, что — если посмотреть на exp[-F/kT], забыв о E — то: exp[-(-TS)/kT] = exp[-(-TklnV)/kT] = V есть просто доступный объем, а он определяет число возможных состояний частицы в пространстве. Чем выше энтропия, тем больше это число состояний, тем вероятнее поэтому нахождение частицы именно в этом объеме.
В общем случае (когда возможные состояния частицы ограничены не только ее столкновениями со стенками, окружающими объем V, но и, скажем, столкновениями c другими частицами), энтропия S частицы равна
S = k´ [логарифм числа доступных для рассматриваемой частицы состояний].
В молекулярной физике, биологии, химии обычно говорят не об одной частице, а о моле (6 x 1023) частиц, и пишут
S = R´ [логарифм числа доступных для одной частицы состояний].
Здесь стоит еще раз напомнить, что различие между k и R — только в том, что k употребляют, говоря об одной частице, а R — о моле частиц.
Величина
F = E — TS | (5.2) |
называется, строго говоря, свободной энергией Гельмгольца. О ней просто рассказывать, и ее просто и удобно вычислять, так как эта величина относится к системе, которая (как та молекула, о которой мы говорили) находится в замкнутом и неизменном объеме.
Однако обычно на опыте измерения ведутся не при постоянном объеме V, а при постоянном внешнем давлении P (например — при атмосферном давлении). В этом случае измеряется не изменение чистой энергии Е изучаемого вещества, а изменение его энтальпии Н=Е+PV: в изменение энтальпии H, помимо собственно изменения энергии тела (Е), входит работа, совершенная против постоянного внешнего давления P при изменении объема V этого тела.
На практике — той, с которой мы будем иметь дело по ходу этих лекций — величину PV можно игнорировать, так как нас будут интересовать жидкости или твердые тела (где на каждую молекулу приходится небольшой объем), причем при невысоком (типа атмосферного) давлении. При этих условиях величина PV много-много меньше, чем тепловая энергия тела.
Поэтому для нас будет в дальнейшем несущественна разница между H и E. Обе эти величины я буду обычно называть просто энергия.
Столь же несущественно для нас будет различие между свободной энергией Гельмгольца F = E — TS и свободной энергией Гиббса
G = Н — TS = (E+PV) — TS = F + PV . | (5.3) |
И то, и другое (и G, и F) мы, не делая различия, будем именовать просто свободной энергией.
Пожалуй, стоит запомнить, что, обсуждая процессы, протекающие в замкнутом объеме, следует употреблять буквы E и F, а обсуждая процессы, протекающие при постоянном внешнем давлении — буквы H и G.
И еще следует запомнить то, что равновесному, стабильному состоянию любой системы при заданной температуре (T=const) соответствует минимум величины F=E - TS, если равновесие устанавливается при фиксированном объеме, и минимум величины G = H- TS, если оно устанавливается при фиксированном внешнем давлении.
При малом изменении состояния системы ее свободная энергия слегка меняется:
F ® F + dF = F + (dE - TdS - SdT) | |
или | (5.4) |
G ® G + dG = G + (dH - TdS - SdT) |
Это значит, в частности, что все возможные перестройки системы в окрестности ее стабильного состояния (а они могут изменять и энергию системы, и ее энтропию) подчиняются следующим уравнениям, характеризующим минимум функции (как мы знаем, точка минимума функции обладает тем свойством, что небольшое отклонение от нее практически не меняет функцию):
1) при V=const стабильному состоянию (при заданном Т) соответствует минимум F, т. е.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
