
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
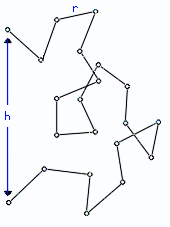
Рис.9-7. Свободно-сочлененная цепь — модель клубка.
Такую цепь можно описать как цепочку из векторов r1, r2 ,..., rM (напомню: вектор в математике — это отрезок прямой, имеющий направление). Эти вектора имеют одинаковую длину r, и каждый из них направлен от предыдущего сочленения к последующему. Сумма векторов
| (9.17) |
есть просто вектор, идущий от начала к концу цепи. Квадрат длины этого вектора есть
| (9.18) |
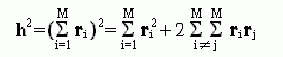
Найдем теперь среднюю величину <h2>, т. е. усредним h2 по всем возможным тепловым флуктуациям цепи. При этом мы должны учесть, что <ri2>, средний квадрат вектора ri, есть просто r2, а среднее произведение любых векторов <rirj> есть 0 при i¹j, так как, свободно вращаясь, эти вектора имеют ту же вероятность быть направленными что в одну, что в разные стороны.
Значит, средний квадрат расстояния между концами свободно-сочлененной цепи есть
| (9.19) |
то есть линейные размеры (радиус и т. д.) клубка растут с ростом числа M звеньев в цепи как M1/2. Отсюда следует, что объем клубка пропорционален M3/2 — в то время как для всех "нормальных" (имеющих фиксированную плотность) тел объем растет только как число частиц M, — т. е. гораздо медленнее, чем M3/2. Такая аномально сильная зависимость объема клубка от размера цепи и есть самое характерное свойство клубка. Из него, в частности, вытекает крайне низкая плотность клубка, образованного длинными цепями, и, как следствие — практическое отсутствие контактов между дальними по цепи звеньями.
Недостаток, недостаточная реалистичность свободно-сочлененной модели — в том, что она рисует клубок в сильно идеализированном виде, т. к. допускает, что в любом своем звене цепь может поворачиваться на любой угол, — а это, вообще говоря, не так. Однако формулу (9.19) можно переписать в более общем виде,
<h2> = Lr, | (9.20) |
где L = Mr — полная ("контурная") длина цепи, пропорциональная числу звеньев в ней, а r — эффективное расстояние между "свободными сочленениями" цепи, т. е. характерная длина, на которой цепь "забывает" о своем направлении (для справки: в полипептидной цепи эта характерная длина — или, как говорят, "длина Куновского сегмента" — составляет 30 — 40![]() , т. е. Куновский сегмент такой цепи содержит около 10 аминокислотных остатков). В виде (9.20) формула, описывающая размер клубка, уже обретает достаточную общность и может применяться для описания реальных полимеров
, т. е. Куновский сегмент такой цепи содержит около 10 аминокислотных остатков). В виде (9.20) формула, описывающая размер клубка, уже обретает достаточную общность и может применяться для описания реальных полимеров
10
Обсудим теперь свойства боковых групп аминокислотных остатков. В особенности я хочу остановиться на том, какие именно структуры стабилизуют те или иные остатки.
Список 20 "стандартных", т. е. кодируемых ДНК аминокислотных остатков дан в Таблице 10/1; там же дан их молекулярный вес и встречаемость в белках. Структуры аминокислотных остатков представлены на картинке 10-1.
Таблица 10/1. Основные свойства аминокислотных остатков
_____ Аминокислотный остаток _____ | % в белках E. coli | Мол вес при pH7 (дальтон) | DGвода®спирт | ||
название | код | ||||
3-букв. | 1-букв. | ||||
Глицин | Gly | G | 8 | 57 | 0 |
Аланин | Ala | A | 13 | 71 | -0.4 |
Пролин | Pro | P | 5 | 97 | -1.0 |
Глутаминовая кислота | Glu | E | »6 | 128 | +0.9 |
Глутамин | Gln | Q | »5 | 128 | +.03 |
Аспарагиновая кислота | Asp | D | »5 | 114 | +1.1 |
Аспарагин | Asn | N | »5 | 114 | +0.8 |
Серин | Ser | S | 6 | 87 | +0.1 |
Гистидин | His | H | 1 | 137 | -0.2 |
Лизин | Lys | K | 7 | 129 | +1.5 |
Аргинин | Arg | R | 5 | 157 | +1.5 |
Треонин | Thr | T | 5 | 101 | -0.3 |
Валин | Val | V | 6 | 99 | -2.4 |
Изолейцин | Ile | I | 4 | 113 | -1.6 |
Лейцин | Leu | L | 8 | 113 | -2.3 |
Метионин | Met | M | 4 | 131 | -1.6 |
Фенилаланин | Phe | F | 3 | 147 | -2.4 |
Тирозин | Tyr | Y | 2 | 163 | -1.3 |
Цистеин | Cys | C | 2 | 103 | -2.1 |
Триптофан | Trp | W | 1 | 186 | -3.0 |
Примечания. Все данные взяты из [3], — за исключением данных по гидрофобности боковых групп, которые взяты из I. I.Fauchere, V. Pliska, Eur. J. Med. Chem.-Chim. Ther. (1983) 18:369. Объем (в ![]() 3), приходящийся на аминокислотный остаток в белке или в растворе, близок к его молекулярному весу в (дальтонах), умноженному на 1.3. Точнее, — процентов на 5 побольше, чем (мол. вес) x 1.3, если в остатке много алифатических (-СН2-, - СН3) групп, и процентов на 5 поменьше, чем (мол. вес) x 1.3, если в остатке много полярных (О, N) атомов.
3), приходящийся на аминокислотный остаток в белке или в растворе, близок к его молекулярному весу в (дальтонах), умноженному на 1.3. Точнее, — процентов на 5 побольше, чем (мол. вес) x 1.3, если в остатке много алифатических (-СН2-, - СН3) групп, и процентов на 5 поменьше, чем (мол. вес) x 1.3, если в остатке много полярных (О, N) атомов.
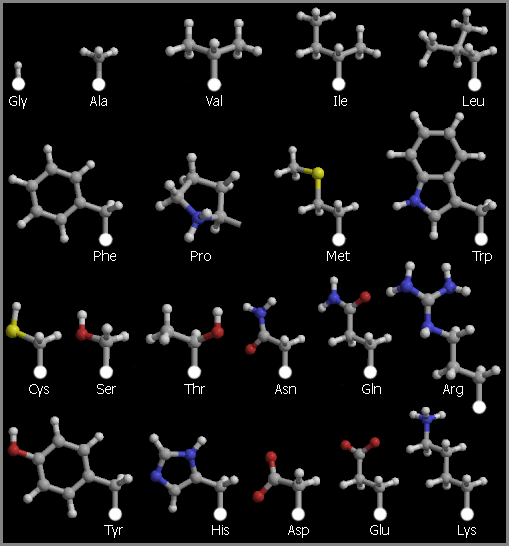
Рис.10-1. Боковые цепи двадцати стандартных аминокислотных остатков.
Рассмотрим теперь структурные тенденции аминокислотных остатков; они стали известными после многолетнего статистического исследования белковых структур. Такие исследования отвечают на вопрос: "Что чаще всего бывает, и чего чаще всего не бывает?".
Для систематизации ответов полезной может быть следующая ниже Таблица 10/2, куда я вписал, наряду со встречаемостью аминокислотных остатков в разных местах белков, такие свойства остатков, как: наличие NH группы в главной цепи (ее нет только у иминокислоты пролина); наличие Сb атома в боковой цепи (его нет только у глицина); число не-водородных g атомов в боковой цепи; наличие и вид полярных группировок в боковой цепи (диполей или зарядов — со знаком; жирным выделено то зарядовое состояние, что относится к "нормальному" рН7).
Таблица 10/2. Основные структурные свойства аминокислотных остатков
A. к. | Наличие | число | Диполь/заряд | pK | Яркая тенденция быть в: | ||||||||
ост. | NH | Cb | g | до | спираль | за | клубок | ядро | |||||
aN | |aN | a | aC| | aC | b | ||||||||
Gly |
| - | - | + | |||||||||
Ala |
|
| + | - | |||||||||
Pro |
| 1 | + | - | - | - | - | + | |||||
Glu |
|
| 1 | COOH Þ CO2- | 4.3 | + | + | - | - | - | - | ||
Asp |
|
| 1 | COOH Þ CO2- | 3.9 | + | + | - | - | - | - | + | - |
Gln |
|
| 1 | OCNH2 | - | ||||||||
Asn |
|
| 1 | OCNH2 | + | - | + | - | + | - | |||
Ser |
|
| 1 | OH | + | + | |||||||
His |
|
| 1 | NH; и N Þ NH+ | 6.5 | - | + | + | |||||
Lys |
|
| 1 | NH2 Þ NH3+ | 10.5 | - | - | + | + | - | - | ||
Arg |
|
| 1 | HNC(NH2)2+ | 12.5 | - | - | + | + | - | + | - | |
Thr |
|
| 2 | OH | + | + | |||||||
Ile |
|
| 2 | + | - | + | |||||||
Val |
|
| 2 | + | - | + | |||||||
Leu |
|
| 1 | + | + | - | + | ||||||
Met |
|
| 1 | + | + | - | + | ||||||
Phe |
|
| 1 | + | - | + | |||||||
Tyr |
|
| 1 | OH Þ O- | 10.1 | - | + | + | |||||
Cys |
|
| 1 | SH Þ S- | 9.2 | - | + | + | |||||
Trp |
|
| 1 | NH | + | + |
Примечания. К "структурным свойствам" отнесена тенденция быть в a-спирали (a), и особо — в ее N - и С-концевых витках, а также — непосредственно перед N - и за С-концом спирали; тенденция быть в b-структуре; тенденция быть в нерегулярных структурах, т. е. "петлях" (включая сюда и b-изгибы цепи); и, наконец, — тенденция быть в гидрофобном ядре глобулы, а не на ее поверхности. Тенденцию "быть" я отмечал значком "+", "не быть " — значком "-". Жирным значком отмечалась особо сильная тенденция.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
