
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
U = [q1q2/eэффr] x exp(-r/D) . | (6.7) |
Третье. Электростатическое взаимодействие — яркий пример не парного взаимодействия частиц (в отличие от, например, Вандерваальсового). Оно зависит не только от расстояния r между зарядами q1 и q2, но и от свойств среды (меняющих и e, и D), и в частности — от расстояний от зарядов до других тел и от формы этих тел (все это влияет на eэфф).
И еще одно добавление. До сих пор я обычно говорил "энергия электростатических взаимодействий". Это говорилось только для простоты слога — как я уже говорил в начале лекции, строго говоря, надо было говорить "свободная энергия".
Более того. Исследуя температурную зависимость электростатических эффектов в водном окружении, можно показать, что энтропийная составляющая в них доминирует, а собственно энергетическая (энтальпийная) составляющая близка к нулю. Это видно из того, что диэлектрическая проницаемость воды меняется от 88 до 55 (т. е. электростатические взаимодействия растут примерно на 40%) при росте абсолютной температуры Т от 273 до 373оК (тоже примерно на 35%). А у взаимодействия, растущего пропорционально абсолютной температуре, есть только энтропийная, но нет энтальпийной части. Значит, в воде весь электростатический эффект связан не с энергией, а с упорядочением воды вокруг зарядов и с изменением этого упорядочения при сближении или отдалении зарядов друг от друга.
Значит, как ни парадоксально, а электростатика в воде имеет энтропийную, а не энергетическую природу, — впрочем, так же, как и гидрофобные взаимодействия или как и образование водородных связей в водном окружении.
Заключая раздел "Элементарные взаимодействия в белках и вокруг", я хочу еще упомянуть о дисульфидных связях и о координационных связях. Не столь массовые, как, например, водородные связи, — эти связи, однако, часто играют важную роль в белках.
Дисульфидные (или SS) связи образуются цистеиновыми (Cys) аминокислотными остатками (боковая группа цистеина: - CbH2-SH). Непосредственное, с выделением водорода (по схеме - CH2-SH + HS-CH2- ® - CH2-S-S-CH2- + H2) окисление цистеинов в белках не идет — оно происходит слишком медленно при комнатной температуре. Однако образование SS связей в белках может быстро происходить при помощи тиол-дисульфидного обмена. Полагают, что в клетке это происходит при участии глутатиона, существующего и в мономерной тиольной (GSH), и в димерной дисульфидной (GSSG) формах, — и происходит по схеме
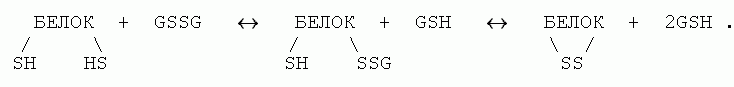
Как разрыв, так и образование SS связей в клетках катализируется (т. е. ускоряется, но не направляется) специальным ферментом — дисульфидизомеразой.
Образование S-S связей в клетке обратимо, так как энергетический баланс этой реакции — тиол - дисульфидного обмена — близок к нулю (было две ковалентные SH связи и одна SS связь, и столько же осталось; не правда ли, это очень походе на энергетический баланс образования внутрибелковых водородных связей в водном окружении?). Более того, имеющаяся — относительно высокая — концентрация GSH в клетке сдвигает равновесие в сторону разрыва тех связей, которые могли бы образоваться при "случайном" сближении цистеинов. Поэтому здесь способны образоваться и выжить SS связи только между "и без того" (т. е. — другими взаимодействиями) сближенными цистеинами.
Особенно важны SS связи для белков, которым придется жить и работать вне клетки. С одной стороны, там нет дисульфидизомераз и глутатиона, так что уже завязавшиеся (внутри клетки или на выходе из нее) связи "замораживаются" — они уже не порвутся и на перестроятся. С другой — белок вне клетки попадает в разные условия, и дополнительный запас прочности, даваемый стабильными замороженными SS связями, ему не помешает. Поэтому SS связи гораздо более типичны для секретированных белков, чем для внутриклеточных. Обычно в секретированных белках все имеющиеся цистеины (кроме одного, если их число нечетно) вовлечены в SS связи.
Координационные связи образуются N, O и S атомами белка (а также О атомами воды) с двух— и трехвалентными ионами металлов: Fe, Zn, Со, Ca, Mg и т. д.
У ионов этих металлов есть незаполненные, но низко лежащие орбитали. Каждая из них способна связать электронную пару. А у О, N, S атомов ("доноров электронов") есть электронные пары, которые могут перетечь на свободные орбитали ионов. Образующаяся при этом связь ничем не отличается от обычной химической связи — кроме того, что для обычной связи электроны поставляются обоими входящими в связь атомами, а для координационной — только одним из них.
При образовании координационных связей ион металла связывается с несколькими донорами электронов. При этом крупные (с радиусом ~1.5![]() ) атомы-доноры со всех сторон окружают маленький (с радиусом ~0.7
) атомы-доноры со всех сторон окружают маленький (с радиусом ~0.7![]() ) двух— или трехвалентный ион. Наиболее часто шесть атомов-доноров окружают (координируют) ион металла, располагаясь по вершинам правильного октаэдра (Рис.6-16).
) двух— или трехвалентный ион. Наиболее часто шесть атомов-доноров окружают (координируют) ион металла, располагаясь по вершинам правильного октаэдра (Рис.6-16).
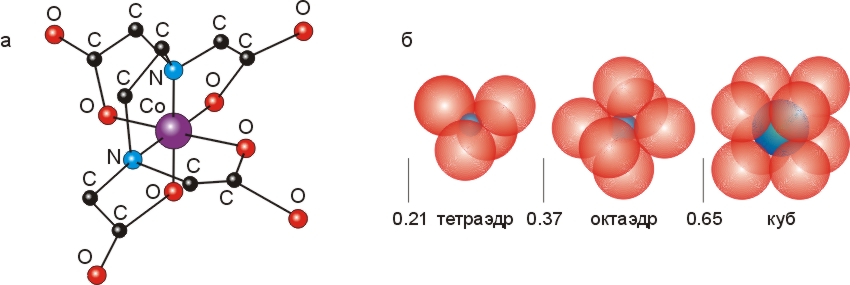
Рис.6-16. (а) Структура октаэдрического комплекса, образованного атомом Со+++ с EDTA. (б) Характерная координация центрального иона при различном соотношении его радиуса с радиусами окружающих его доноров электронов. Картинки взяты из [10] и адаптированы.
Так как ион может связаться и с электронными донорами белка, и с кислородами воды, он (несмотря на большую энергию каждой связи) переходит из воды на белок и обратно без драматического выигрыша или проигрыша энергии. Важнее, пожалуй, другое: если в белке атомы - доноры уже стоят в "правильных" (для образования координационной связи) положениях, — ион может связаться с ними, отпустив ранее окружавшие его молекулы воды, а это приведет к прочной связи из-за выигрыша энтропии движения отпущенных молекул воды. В среднем, каждая координационная связь стоит несколько ккал/моль — несколько больше, чем водородная связь в воде.
Такие связи, образуемые несколькими атомами одной молекулы, способными связать один ион, называются хелатными ("клешневидными").
Роль таких связей в белках, и в частности в их активных центрах, мы рассмотрим позже. Мы увидим также, что хелатные, полностью обволакивающие ион комплексы могут входить в гидрофобное ядро белка. А пока я хочу снова обратить ваше внимание на Рис.6-16, где изображен часто используемый в лабораторной практике реагент ЭДТА (этилендиаминтетраацетат) в хелатной связи с металлом. У ЭДТА эта связь особенно прочна из-за связи отрицательных зарядов СОО- групп ЭДТА с положительно заряженным ионом металла.
Лекция 7
Разобравшись с элементарными взаимодействиями, — рассмотрим сегодня вторичную структуру белков. Прежде всего у нас речь пойдет о регулярных вторичных структурах — об a-спиралях и о b-структуре, — но не только о них.
Укладка a и b-структур в глобулу определяет третичную структуру белка (Рис.7-1). Эти вторичные структуры отличаются определенными, периодическими конформациями главной цепи — при разнообразии конформаций боковых групп.
Рис.7-1. Вторичная структура полипептидной цепи (a-спираль и тяж b-листа) и третичная структура белковой глобулы.
Начнем со спиралей. Они могут быть левые и правые (Рис.7-2), у них может быть разный период и шаг. Правые (R) спирали приходят к нам, завиваясь против часовой стрелки (что отвечает положительному отсчету угла в тригонометрии); левые (L) — приходят, вращаясь по стрелке.
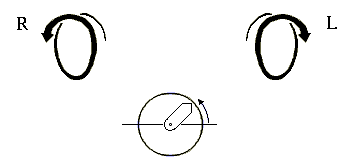
Рис.7-2. Правые (R) и левые (L) спирали. Под ними показан отсчет положительного угла в тригонометрии: при этом "близкая к нам" стрелка вращается против хода часов.
Важнейшие спирали в полипептидной цепи держатся водородными связями, где С=О группы остова полипептида связаны с лежащими от них в направлении С-конца цепи H-N группами. В принципе, возможны следующие спирали, стянутые Н-связями (Рис.7-3): 27, 310, 413 (обычно именуемая a) и 516 (она же p). Здесь в названии "27" — "2" означает связь со 2-м по цепи остатком (см. Рис.7-3), а "7" — число атомов в цикле (O......H-N-C'-Ca-N-C'), замыкаемом этой связью. Тот же смысл имеют цифры и в названии других спиралей.
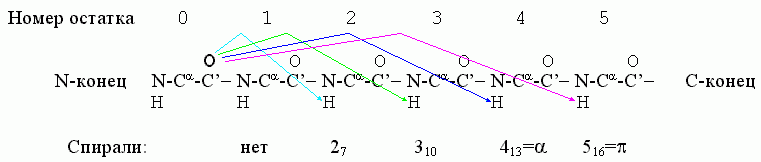
Рис.7-3. Водородные связи (они показаны стрелками), характерные для разных спиралей.
Какие из этих спиральных структур преобладают в белках? a-спирали. Почему? Ответ на этот вопрос дает карта Рамачандрана для типичного аминокислотного остатка — аланина (Рис.7- 4), на которой отмечены конформации, периодическое повторение которых приводит к завязыванию изображенных на Рис.7-3 водородных связей.
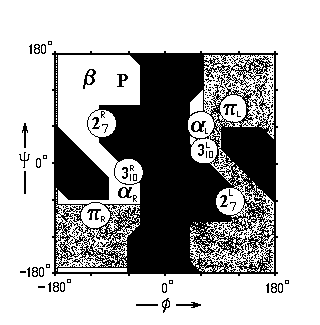
Рис.7-4. Конформации различных вторичных структур на фоне карты разрешенных и запрещенных конформаций аминокислотных остатков. 27R, 27L: правая и левая спираль 27; 310R, 310L: правая и левая спираль 310; aR, aL — правая и левая a-спираль; pR, pL — правая и левая p-спираль. b — b-структура (подробности см. на Рис.7-8б). Р — спираль Poly(Pro)II. ![]() — конформации, разрешенные для аланина (Ala);
— конформации, разрешенные для аланина (Ala); ![]() — области, разрешенные лишь для глицина, но не для аланина и других остатков;
— области, разрешенные лишь для глицина, но не для аланина и других остатков; ![]() — области, запрещенные для всех остатков. j и y — углы внутреннего вращения в белковой цепи.
— области, запрещенные для всех остатков. j и y — углы внутреннего вращения в белковой цепи.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
