
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Видно, что только спираль aR (a-правая) лежит достаточно глубоко внутри области, разрешенной для аланина (и для всех других остатков). Другие спирали лежат либо на краю этой области (например, левая спираль aL или правая спираль 310), где конформационные напряжения уже возрастают, либо в области, доступной только глицину.
Поэтому можно ожидать, что именно правая a-спираль должна быть, как правило, более стабильной, и потому преобладать в белках — что и наблюдается. В правой a-спирали (Рис.7-5) все атомы упакованы оптимально: плотно, но без напряжений; поэтому не удивительно, что в белках таких спиралей много, а в фибриллярных белках они достигают гигантской длины и включают сотни аминокислотных остатков.
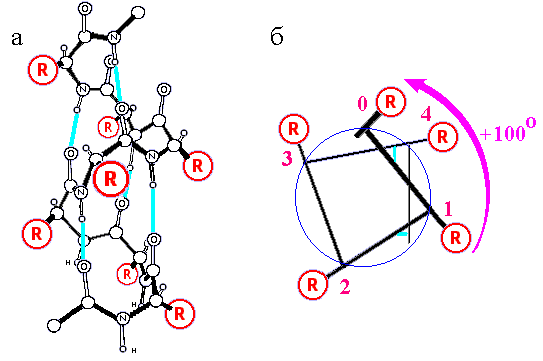
Рис.7-5. Правая a-спираль. (а) Атомарная структура. R — боковые группы. Голубые линии — водородные связи. (б) Схематическое изображение одного витка той же a-спирали, вид с торца. Стрелка показывает поворот спирали (в расчете на один остаток) по мере ее приближения к нам. Картинка (а) взята из [3] и адаптирована.
Левых a-спиралей в белках практически нет. Нет и спиралей 27, которые, мало того что лежат на самом краю разрешенной области, но еще имеют энергетически невыгодный, почти прямой угол схождения N-H и О=С групп.
Практически нет в белках и спиралей p. Они тоже лежат на самом краю разрешенной области, да еще и витки в них слишком широки, так что p-спирали имеют энергетически невыгодную пустую "дырку" на оси. А вот спирали 310 (в основном — правые, — левые пригодны практически лишь для глицинов) в белках есть — правда, в виде коротких (из трех-четырех остатков) и деформированных фрагментов (из-за стерических напряжений: слишком тугая спираль! — соответствующая ей конформация лежит на самом краю разрешенной области).
Отметим одно свойство спиралей, хорошо видное на Рис.7-5а: на их N-конце сидят свободные от внутриспиральных водородных связей Н атомы N-H групп, а на С-конце — свободные от водородных связей О атомы С=О групп. Так как электронное облако с Н атома частично стянуто электроотрицательным N атомом, а электроотрицательный О атом сам стягивает электрон с С' атома, — N-конец спирали несет положительный, а С-конец — отрицательный парциальный заряд. То есть спираль представляет собой длинный диполь: величина суммарного (поставляемого тремя NH-группами) парциального "+" заряда на ее N-конце составляет около половины протонного, а "-" заряда на С-конце a-спирали — около половины электронного заряда.
Теперь рассмотрим регулярные структуры без водородных связей внутри каждой, но соединенные водородными связями между собой.
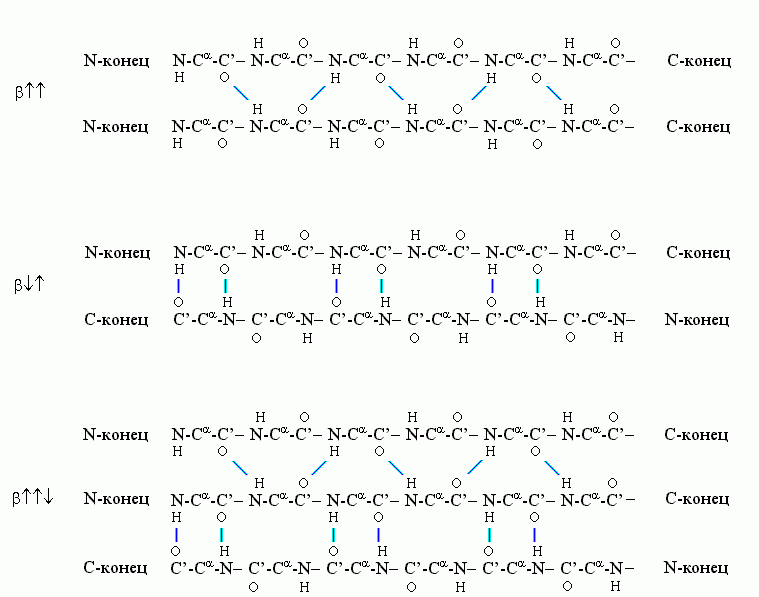
Рис.7-6. Схема хода цепи и расположения водородных связей в параллельной b![]()
![]() , антипараллельной b
, антипараллельной b
 и смешанной b
и смешанной b![]()
![]() b-структуре. Видно, что Н-связи одного остатка в b-тяже направлены одну сторону, следующего — в противоположную, и т. д.
b-структуре. Видно, что Н-связи одного остатка в b-тяже направлены одну сторону, следующего — в противоположную, и т. д.
Почти вытянутые (все углы в главной цепи — почти trans), слегка скрученные цепи образуют b-структуру. Она бывает (Рис.7-6) параллельной (b![]()
![]() ), антипараллельной (b) и смешанной (состоящей из b
), антипараллельной (b) и смешанной (состоящей из b![]()
![]() и b
и b![]()
![]() ).
).
b-структура стянута водородными связями (символы / , \ и | на схемах 7-6). Она существует в виде более или менее крупных листов. Так как поверхность b структуры — рифленая, ее еще называют "складчатой b-структурой" (Рис.7-7).
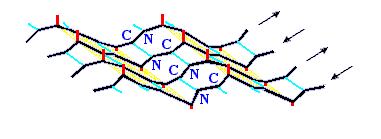
Рис.7-7. Лист b структуры имеет складчатую поверхность. Боковые группы (см. маленькие отростки) расположены на складках; каждая обращена в ту же, что и складка, сторону, т. е. направленные вниз и вверх боковые группы чередуются вдоль b-тяжа. Картинка взята из [3] и адаптирована.
b-структурные листы всегда несколько скручены (Рис.7-8а) из-за того, что несколько скручены (Рис.7-8б) отдельные b-тяжи — и потому по ходу тяжа несколько меняется направленность водородных связей. А тяжи, в свою очередь, скручены из-за того, что наиболее энергетически выгодная конформация остатков с боковыми группами сдвинута к центру стерически разрешенной области (Рис.7-8в). Скрученность отдельного b-тяжа — левая (у L аминокислот! — у D было бы наоборот): вы видите (Рис.7-8б), что боковые группы тяжа поворачиваются по часовой стрелке (на »-165o на каждый остаток) по мере приближения тяжа к нам.
В результате, поворачиваются и Н-связи (на »-165o на каждый остаток, т. е. на -330o=+30o на пару остатков - повторяющуюся единицу b-структуры), так что угол между соседними тяжами b-структуры (если смотреть с кромки листа, Рис.7-8а) обычно составляет около -25о ("минус", как всегда, означает, что ближний к нам b-тяж повернут по часовой стрелки относительно более далекого). Таким образом, b-лист имеет левое скручивание, если смотреть с края этого листа (и правое, если смотреть вдоль b-тяжей на поворот линии Н-связей).
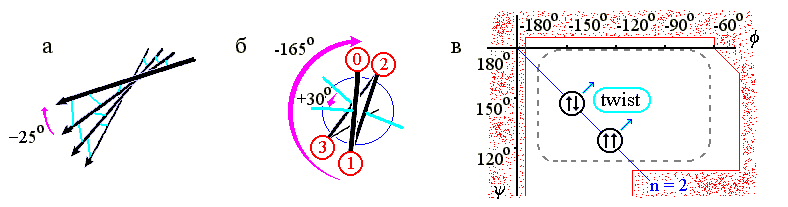
Рис.7-8. (а) Скрученность b-листа. b-тяжи изображены стрелками, водородные связи между ними — голубыми линиями. (б) Схематическое изображение одного витка b-тяжа, вид с торца. Кружки — боковые группы; их номера возрастают по мере удаления от читателя. Голубые линии укаывают направление С=О групп, завязывающих Н-связи в листе, большей стрелкой — поворот b-тяжа при приближении к нам на один остаток, меньшей стрелкой — поворот направленных в одну сторону водородных связей при приближении к нам на два остатка. (в) Конформация идеальной (не скрученной) параллельной (![]()
![]() ) и антипараллельной (
) и антипараллельной (![]()
![]() ) b-структуры для поли(Gly), и усредненная конформация реальной (сложенной из L аминокислот) скрученной (twist) b-структуры. Пунктир показывает область энергетического минимума для отдельно взятого остатка Ala; контуром показаны границы области разрешенных конформаций этого остатка. Диагональ jy-карты соответствует плоской периодичной структуре, имеющей 2 остатка на виток. Над диагональю лежат левые (L) спирали, под ней — правые (R). Картинки (а), (в) взяты из [3] и адаптированы.
) b-структуры для поли(Gly), и усредненная конформация реальной (сложенной из L аминокислот) скрученной (twist) b-структуры. Пунктир показывает область энергетического минимума для отдельно взятого остатка Ala; контуром показаны границы области разрешенных конформаций этого остатка. Диагональ jy-карты соответствует плоской периодичной структуре, имеющей 2 остатка на виток. Над диагональю лежат левые (L) спирали, под ней — правые (R). Картинки (а), (в) взяты из [3] и адаптированы.
Есть спирали и без водородных связей, где плотная (а значит — энергетически выгодная) упаковка держится чисто на Вандерваальсовых контактах. Это — полипролиновая спираль. При этом три скрученные в довольно растянутую левую спираль цепи образуют правую суперсуперспираль — они плотно закручиваются друг вокруг друга. Из двух возможных типов полипролиновой спирали для нас важна спираль poly(Pro)II: она реализуется в коллагене. В этой спирали пептидные группы пролинов находятся в обычной (trans) конформации. Отложим более подробное рассмотрение коллагеновой спирали до соответствующего места курса, а пока ограничимся общим ее видом (Рис.7-9) и отметим на Рис.7-4 область соответствующей ей конформации цепи: видно, что она довольно близка к b структуре.
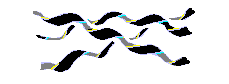
Рис.7-9. Общий вид тройной правой суперспирали из левых спиралей Poly(Pro)II.
Параметры наиболее важных регулярных вторичных структур белковых цепей суммируются в следующей таблице:
Таблица 7/1. Основные геометрические параметры наиболее распространенных в белках вторичных структур.
Структура | H-связь | Остаток/виток | Смещение/остаток ( | f | y |
Спираль aR | CO0—HN+4 | +3.6 | 1.5 | -600 | -450 |
Спираль (310)R | CO0—HN+3 | +3 | 2.0 | -500 | -250 |
Лист b | меж цепей* | -2.3 | 3.4 | -1350 | +1500 |
Лист b | меж цепей* | -2.3 | 3.2 | -1200 | +1350 |
Спираль Poly(Pro) II | нет | -3 | 3.0 | -800 | +1550 |
* Расстояние между тяжами в b-листе: 4.8![]()

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
