
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
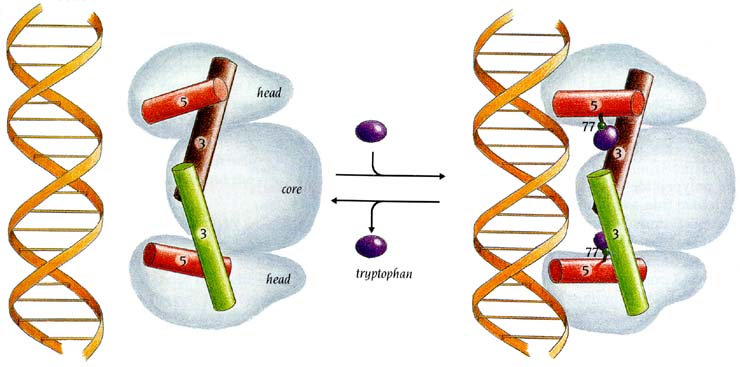
Рис.20-3. Схема действия триптофанового (trp) репрессора. На фоне общего контура димера, обладающего общим сплавленным ядром (core) и двумя идентичными головками (head), показаны только те две спирали (3 и 5), между которыми садится корепрессор — аминокислота Trp. Только при этом обе спирали 5 могут связаться с ДНК. Картинка взята из [5].
Триптофан в данном случае выступает стимулятором ДНК-связывающей активности trp-репрессора и ингибитором синтеза белков, необходимых для синтеза триптофана. Причем это стимулирование trp-репрессора является "аллостерическим", т. к. Trp связывается с белком "в другом месте", — не в том, где с белком должна связываться ДНК.
Мотив "спираль-изгиб-спираль", фигурирующий на Рис.20-1 и Рис.20-3 — характерный, но отнюдь не единственный структурный мотив, связывающий ДНК. Чтобы подчеркнуть это, я вынес три других характерных мотива на Рис.20-4. Я хотел подчеркнуть, что ДНК-связывающие белки могут принадлежать к разным структурным классам (на представленных рисунках есть и a, и a+b белки), и что даже само связывание с ДНК может осуществляться как a-, так и b-структурой.
а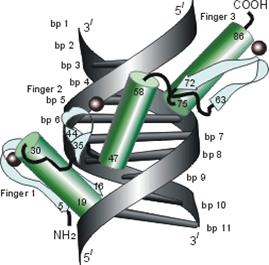 б
б 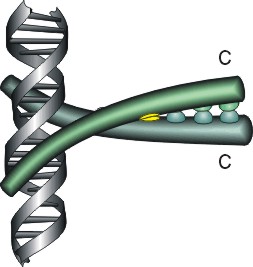 в
в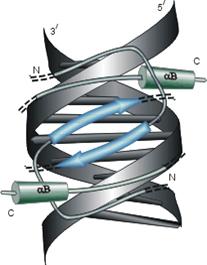
Рис.20-4. Еще три характерных ДНК-связывающих белковых мотива. В двух из них ключевая роль принадлежит a-спиралям: (а) "цинковые пальцы" (шарики — ионы Zn) и (б) "лейциновый зиппер". В третьем, met-репрессоре (в) — ключевая роль принадлежит b-шпильке: она специфически связывается с большим желобком ДНК, в то время как a-спирали aВ связываются неспецифически с сахаро-фосфатным остовом ДНК. Цинковый палец, "finger" (этот домен можно отрезать от целого белка и выделить отдельно) — самый маленький из известных глобулярных белков, а лейциновый зиппер — самый простой из них по структуре. Когда лейциновый зиппер не связан с ДНК, он представляет собой просто димер из параллельных a-спиралей, слепившихся по всей длине своими узкими гидрофобными поверхностями. Образующие эти поверхности боковые группы показаны в виде выступов. Однако одна (на каждой спирали) из этих групп — не гидрофобна: желтым пятном отмечен вкрапленный в гидрофобную поверхность полярный Asn. Он необходим для образования именно димера, т. к. его замена на более гидрофобный остаток приводит к тому, что спирали слипаются не по две, а по три и более. Картинки, с небольшими изменениями, взяты из [6] (а) и [5] (б, в). У каждой цепи помечен ее N - и/или С-конец.
До сих пор мы говорили о тех грубых (характерный размер: ~10-30 ![]() ) чертах белковой структуры, которые позволяют ей внедриться в желобок ДНК. За опознание же той конкретной ДНКовой последовательности, с которой белок должен связаться, отвечают более мелкие черты белковой поверхности (характерный масштаб — размер атома, ~3
) чертах белковой структуры, которые позволяют ей внедриться в желобок ДНК. За опознание же той конкретной ДНКовой последовательности, с которой белок должен связаться, отвечают более мелкие черты белковой поверхности (характерный масштаб — размер атома, ~3 ![]() ).
).
К сожалению, "общий код" выборочного опознавания белками фрагментов ДНК пока не выяснен (и не ясно, есть ли такой сколько-нибудь четкий "код"), — хотя, глядя на детали каждого расшифрованного ДНК-белкового контакта, можно увидеть, какие водородные связи между боковыми группами белка и нуклеотидами, и какие их другие плотные контакты способствовали образованию ДНК-белкового контакта именно в этом месте.
Тонкое, выборочное опознавание белками других молекул удобно рассмотреть на примере иммуноглобинов, или антител — белков, предназначенных (в организмах позвоночных) для тонкого распознавания мелкомасштабных (размером в атом или несколько атомов) антигенных детерминант у самых разных молекул самого разного размера. А аналогичные иммуноглобинам рецепторы Т-клеток опознают таким же образом маленькие антигенные детерминанты у клеток, — например, у клеток, зараженных вирусами.
Иммуноглобины состоят из многих b-структурных доменов и относительно небольших гибких шарниров между ними (Рис.20-5а). Разнообразие сочетаний вариабельных (антиген-связывающих) доменов обеспечивает иммуноглобинам широкий спектр сортов и, соответственно, широкий спектр действия, а твердость этих доменов — высокую селективность действия иммуноглобина каждого сорта. Я не буду пересказывать основы клонально-селекционной теории происхождения огромного разнообразия иммуноглобинов. [Вы должны помнить из других курсов, что в зародышевых клетках представлены не целые гены легких и тяжелых цепей иммуноглобинов, а части, куски этих генов. Там эти части собраны в кассеты, — отдельно много сортов для каждого из трех кусков вариабельного домена тяжелой цепи, отдельно — легкой цепи; отдельно — константные домены каждой цепи, отдельно — шарниры. При образовании соматических иммунных клеток эти куски всячески тасуются — и еще каким-то загадочным образом мутируют в своих гипервариабельных участках — и соединяются в целые гены легких и тяжелых иммуноглобиновых цепей.]
а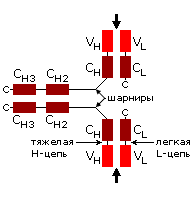 б
б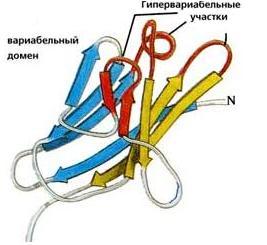 в
в г
г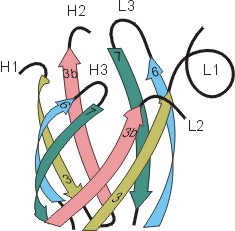
Рис.20-5. (а) Общее строение одного из иммуноглобинов (IgG). Отмечены вариабельные (V) и константные (С) домены двух легких (L: из двух доменов каждая) и двух тяжелых (H: из четырех доменов каждая) цепей, а также С-концы всех этих цепей. Домены СН2 гликозилированы. Жирные стрелки указывают на антиген-связывающие "карманы" между доменами VL и VH. (б) Строение домена VН; почти так же выглядят домены VL, СL, СН и прочие. В показанном домене VH выделены гипервариабельные петли; гипервариабельные петли есть и в домене VL. Вместе эти два домена образуют антиген-связывающий карман (в). (г) Антипараллельный b-цилиндр, образованный b-листом домена VH и b-листом домена VL. Антиген-связывающий карман образуют исходящие из этих листов гипервариабельные петли тяжелой и легкой цепи (Н1, Н2, Н3 и L1, L2, L3, соответственно). Картинки (б, в, г) взяты из [5].
Для нас важно сейчас, что антиген опознается вариабельными доменами легкой и тяжелой цепи (VH и VL) совместно, — точнее, их (VH и VL) гипервариабельными петлями, оторачивающими антиген-связывающий карман, находящийся на стыке этих двух доменов (Рис.20-5,б-г). Первичная структура этих петель варьирует от одного сорта молекул иммуноглобина к другому (что и создает огромное разнообразие этих сортов), но, для каждого данного сорта, — не только аминокислотная последовательность, но и конформация всех петель строго фиксирована, а сам антиген-связывающий карман покоится на твердом b-цилиндре, образованном соединившимися в рукопожатии антипараллельными b-листами вариабельных доменов. Поэтому каждая молекула иммуноглобина может сильно связать только определенный антигенный детерминант — и равнодушна к другим.
Рисунок 20-6 показывает, что селективность связывания антигенных детерминант определяется не устройством белка в целом (оно служит лишь как бы фундаментом), а прежде всего комплементарностью формы, обводов связываемой молекулы к форме относительно небольшой вмятины, к форме только самого антиген-связывающего кармана.
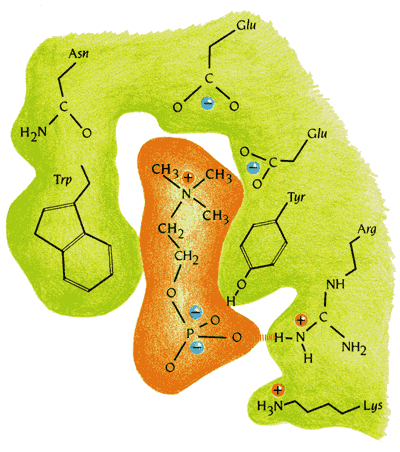
Рис.20-6. Специфическое взаимодействие антигена и связывающего его "кармана" антитела. Показаны сближающиеся заряды и образующиеся водородные связи. Картинка взята из [5].
Кроме того, гидрофобные части связываемой молекулы контактируют с гидрофобными частями кармана, его заряды комплементарны зарядам, вкрапленным в карман, а находящиеся на антигене доноры и акцепторы водородных связей комплементарны вкрапленным в карман антитела акцепторам и донорам этих связей. Все это делает связывание — но только связывание строго определенного антигена — крепким.
Такое же, как в антителах, расположение активного центра — в воронке на торце b-цилиндра — наблюдается и во многих других белках, никак с иммуноглобинами не связанных. Например, это — стандартное место активного центра в a/b цилиндрах (где, в отличие от иммуноглобинов, b-структура параллельна). Вообще, изучая структуры белков, легко заметить, что очень часто активный центр помещается в их "стандартном дефекте", в стандартно расположенной (т. е. определяемой мотивом укладки цепи, а не боковыми группами) вмятине в архитектуре белковой глобулы (Рис.20-7): такая вмятина автоматически способствует окружению субстрата одновременно многими боковыми цепями белка.
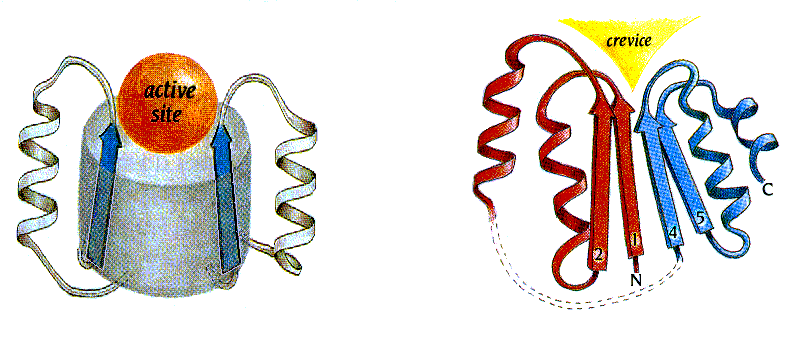
Рис.20-7. Стандартные вмятины в архитектурах белковых глобул часто определяют место-положение (не функцию!) активного центра. Слева: активный центр (active site) в воронке при верхушке b/a-бочонка с параллельным b-цилиндром; сходное расположение активного центра в воронке при верхушке антипараллельного b-цилиндра смотри на Рис.20-5г. Справа: активный центр в щели (crevice), образующейся в укладке Россманна в месте расхождения правых b-a-b суперспиралей, идущих в разные стороны (в суперспирали b1-a-b2 цепь идет от нас, в суперспирали b4-a-b5 — к нам). Картинки взяты из [5].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
