
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
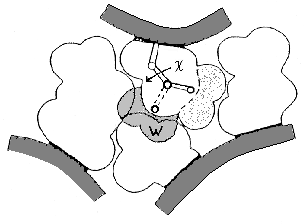
Рис.16-11. Схема упаковки боковых групп. Показан лишь маленький фрагмент ядра. Заштрихованная область W соответствует альтернативному ротамеру боковой группы (c — угол ее вращения); этот ротамер запрещен плотной упаковкой. Для его появления нужен дополнительный объем W » 30 |
|
| Рис.16-12. Причина существования свободно-энергетического барьера между нативным и любым денатурированным состоянием белка. "Барьерное" состояние возникает при небольшом расширении нативного. В этом состоянии поры в белке уже приводят к значительному повышению Вандерваальсовой энергии, но еще не позволяют ни поворотной изомеризации, ни проникновения воды ( |
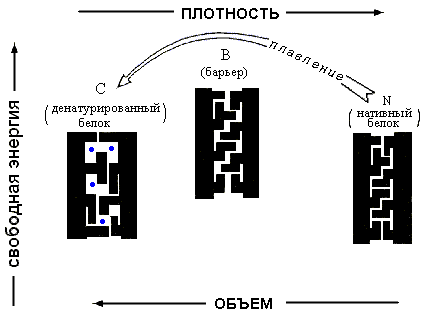
Поэтому малое ("до-барьерное") расширение нативной глобулы всегда невыгодно — оно повышает ее энергию (т. к. части глобулы уже расходятся), но не повышает ее энтропию, так как еще не открывает поворотной изомеризации ее боковых групп. То есть свободная энергия глобулы растет при малом расширении. Наоборот, большое ("за-барьерное") расширение глобулы открывает поворотную изомеризацию и ведет (при достаточно высокой температуре) к падению ее свободной энергии. В результате денатурация белка при изменении внешних условий происходит не постепенно, а скачком, — по принципу "все или ничего".
То есть белок, не меняясь, терпит изменение внешних условий до некоторого предела, — а потом плавится, как микроскопическое твердое тело, весь сразу. Такая устойчивость и твердость белка, в свою очередь, обеспечивает надежность его работы в организме.
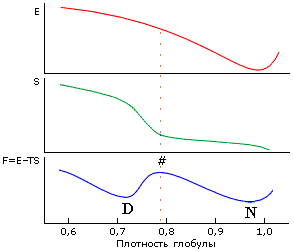
Иными словами, фазовый переход между нативным и денатурированным состояниями объясняется скачкообразным ростом энтропии (и прежде всего — энтропии боковых групп) при расширении глобулы (Рис.16-13), а его фазовый, кооперативный характер связан с тем, что боковые группы прикреплены к главной цепи и не могут раскрепощаться поодиночке.
Рис.16-13. Причина наличия перехода "все-или-ничего" при денатурации белка. Энергия Е минимальна при плотной упаковке глобулы (здесь ее плотность r = 1) и монотонно растет при сжатии или расширении. Энтропия S растет с падением плотности r сперва медленно (при r = 1.0 — 0.8, пока боковые группы лишь колеблются, а их поворотная изомеризация еще невозможна), затем быстро (когда поворотная изомеризация начинает раскрепощаться), и затем (когда поворотная изомеризация уже свободна) — опять медленно. Неравномерный рост энтропии связан со следующим эффектом. Для поворотной изомеризации вблизи боковой группы должен быть свободный объем размером по крайней мере с СН3-группу. А так как боковая группа привязана к жесткой главной цепи, такой объем не может появиться у одной лишь группы, — он одновременно появится у многих. Поэтому и существует характерная ("барьерная") плотность глобулы, где энтропия начинает резко расти с расширением глобулы. Эта плотность довольно высока: »80% от нативной плотности, т. к. объем СН3-группы (характерной поворачивающейся единицы, см. Рис.16-11) составляет »20% объема аминокислотного остатка. В результате зависимость суммарной свободной энергии F=E-TS от плотности глобулы проходит через максимум ("барьер"), отделяющий нативную (плотноупакованную) глобулу (N) от любого более рыхлого денатурированного состояния (D). Из-за этого барьера денатурация белка и происходит по типу "все или ничего", независимо от дальнейшего состояния денатурированной молекулы. |
|
До открытия расплавленной глобулы денатурация белков обычно трактовалась как полное разрушение структуры и переход глобулы в клубок. После этого открытия стало ясно, что денатурированный белок может быть как довольно плотным, так и рыхлым — в зависимости от силы растворителя и гидрофобности цепи. Оценки показывают, что поры расплавленной глобулы (поры, необходимые для свободы движений боковых групп) заполнены растворителем, так как перенос молекулы воды в п![]() ру внутри белка термодинамически выгоден по сравнению с существовавшим там до того вакуумом. На опыте об этом свидетельствует ничтожность изменения парциального объема при денатурации белка. Но проникающему внутрь растворителю, если только он не слишком сильно притягивается п
ру внутри белка термодинамически выгоден по сравнению с существовавшим там до того вакуумом. На опыте об этом свидетельствует ничтожность изменения парциального объема при денатурации белка. Но проникающему внутрь растворителю, если только он не слишком сильно притягивается п![]() рами (ведь они окружены в основном неполярными группами, слабо притягивающими воду), — ему удастся только заполнить поры уже существующие, но не удастся создать новые, не удастся разорвать оставшиеся в белке взаимодействия. Тогда он не сможет расширить глобулу — как вода не может растворить губку, хоть и заполняет ее поры, — и белок так и останется компактной расплавленной глобулой (причем "мокрой", насыщенной водой расплавленной глобулой). Компактность такой глобулы обеспечивается остаточными гидрофобными взаимодействиями. Если же растворитель сильно притягивается к порам (т. е. если или растворитель сильнее, или поры менее гидрофобны), то он начинает расширять поры, глобула начнет разбухать, — и тем сильнее, чем сильнее притяжение растворителя к белковой цепи, т. е. чем меньше выгоды находят звенья цепи в контакте друг с другом. По мере роста притяжения растворителя к белковой цепи она разбухнет до неупорядоченного клубка, и этот процесс как раз и описывается теорией переходов глобула-клубок, иллюстрируемой Рис.16-10.
рами (ведь они окружены в основном неполярными группами, слабо притягивающими воду), — ему удастся только заполнить поры уже существующие, но не удастся создать новые, не удастся разорвать оставшиеся в белке взаимодействия. Тогда он не сможет расширить глобулу — как вода не может растворить губку, хоть и заполняет ее поры, — и белок так и останется компактной расплавленной глобулой (причем "мокрой", насыщенной водой расплавленной глобулой). Компактность такой глобулы обеспечивается остаточными гидрофобными взаимодействиями. Если же растворитель сильно притягивается к порам (т. е. если или растворитель сильнее, или поры менее гидрофобны), то он начинает расширять поры, глобула начнет разбухать, — и тем сильнее, чем сильнее притяжение растворителя к белковой цепи, т. е. чем меньше выгоды находят звенья цепи в контакте друг с другом. По мере роста притяжения растворителя к белковой цепи она разбухнет до неупорядоченного клубка, и этот процесс как раз и описывается теорией переходов глобула-клубок, иллюстрируемой Рис.16-10.
Иными словами: если белковая цепь достаточно гидрофобна, денатурированный теплом белок останется в состоянии расплавленной глобулы; если недостаточно — он развернется.
Разворачивание расплавленной глобулы выглядит как кооперативный переход (Рис.16-8). Однако — является ли это разворачивание переходом типа "все-или-ничего"?
Здесь пока нет полной ясности. Термодинамические исследования многих белков показывают, что, по всей видимости, — нет, не является. Однако недавно Уверским и Птицыным было показано, что состояние расплавленной глобулы может быть, по крайней мере в некоторых белках, отделено от "более развернутого" состояния переходом типа "все-или-ничего", и что только дальнейшее разворачивание этого "более развернутого" состояния до клубка является плавным, как того требует показанная на Рис.16-10 теория.
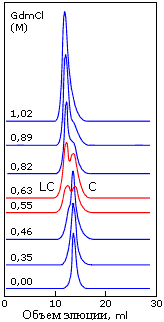
Сосуществование двух фаз — "расплавленных" и "менее плотных" глобул — наблюдается при хроматографировании белка в области таких концентраций денатуранта, где расплавленная глобула разваливается. Наличие раздвоенного пика на таких хроматограммах (Рис.16-14) доказывает, что данный белок существует либо в одной форме (расплавленной — это установлено другими измерениями), либо в другой (какой-то "менее плотной") — но не в форме глобулы какой-то промежуточной между ними плотности.
| Рис.16-14. Сосуществование двух фаз, "компактных расплавленных глобул" (С) и "менее плотных глобул" (LC) при хроматографировании белка в области таких концентраций денатуранта (гуанидингидрохлорида), где расплавленная глобула начинает разваливаться. Обратите внимание на раздвоенность пика на хроматограммах, сделанных при 0.55 — 0.63 М GdmCl. Картинка взята из V. N.Uversky, G. V.Semisotnov, R. H.Pain & O. B.Ptitsyn, FEBS Letters (1992) 314:89-92. |
Итак, возвращаясь к фазовой диаграмме Танфорда (Рис.16-6), мы видим, что нативное состояние отделено от всех других (и от клубка RC, от температурно-денатурированного состояния НD) переходом типа "все-или-ничего" (что и обуславливает сравнительную узость этих переходов); что денатурированное состояние обязательно отличается от нативного подвижными боковыми группами, но может быть весьма компактным; и, наконец, что переход из температурно-денатурированного состояния в клубок может быть как фазовым переходом типа "все-или-ничего" для одних белков, так и (возможно) не-фазовым переходом — для других.
Маленькое отступление. Вообще, нас не должно удивлять, что разные белки ведут себя по-разному. И здесь дело не столько в качественных различиях между белками, сколько в том, что мы наблюдаем каждый белок через "экспериментальное окошко", где температурный диапазон ограничен существованием жидкой воды, а диапазон воздействия денатуранта — его нулевой концентрацией. Поэтому, например, менее гидрофобные по своему аминокислотному составу белки — белки, легко разрушаемые денатурантом — будут демонстрировать в этом "окошке" только переход в клубок (ведь расплавленная глобула держится только остаточными гидрофобными взаимодействиями). И эти же малостабильные белки (или белки "ослабленные" — как, например, апомиоглобин, т. е. миоглобин, ослабленный лишением гема), — эти белки демонстрируют холодовую денатурацию — денатурацию, при которой всегда образуется клубок. У более стабильных белков она не наблюдается — точнее, она, видимо, просто приходится на температуры, когда вода замерзает и тем прекращает наши опыты...

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
