
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Тогда обратились к маленьким (из 50-100 остатков) белкам, сворачивающимся in vitro без наблюдаемых, "накапливающихся" в эксперименте интермедиатов, и начали исследовать их более интенсивно. И первая неожиданность, с которой здесь столкнулись, — оказалось, что эти белки порой сворачиваются очень быстро — много быстрее тех, наличие интермедиатов в сворачивании которых должно было, казалось бы, ускорить и облегчить их правильное сворачивание!
Обычно белки "однодоменного" размера сворачивались за секунды или десятки секунд, но некоторые из этих белков (правда, это были маленькие белки — до 100 остатков длиной) сворачивались — полностью, до нативной структуры, без наблюдаемых накапливающихся промежуточных структур — за миллисекунды (Рис.17-4). Это регистрировалось по одинаковой скорости восстановления вторичной структуры, упаковки боковых групп, флюоресценции триптофанов (зависящей от их погруженности в глобулу), обмениваемости водородов в белке и т. д.
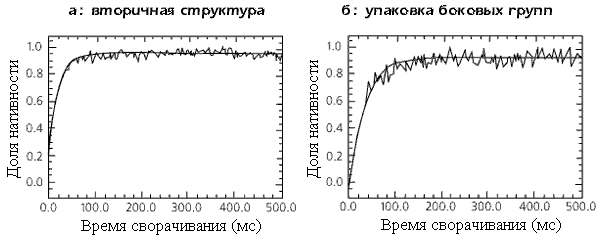
Рис.17-4. Ренатурация ACBP (белка, связывающего ацил-коэнзим А), регистрируемая по восстановлению эллиптичности при 225 нм (а) и 286 нм (б). Тонкая, ломаная линия — экспериментальный сигнал, жирная кривая — результат его сглаживания, с учетом погрешности опыта. Картинки взяты из Kragelund B. B., Robinson C. V., Knudsen J., Dobson C. M. & Poulsen F. M. Biochemistry (1995) 34: 7217-7224.
Однако, — что в таких экспериментах можно исследовать, чтобы пролить свет на природу процесса самоорганизации? Ведь интермедиатов, которые можно выделить и изучать, тут нет?!
Ответ: здесь можно изучать переходное состояние — состояние, играющее ключевую роль в кинетике процесса.
Напомню, что кинетика перехода между двумя непосредственно наблюдаемыми состояниями (А и В на Рис.17-5) успешно описывается теорией переходных состояний — которая в химии обычно называется теорией активированных комплексов. Суть ее сводится к тому, что скорость процесса лимитируется переходным состоянием #, т. е. наименее (по термодинамике) вероятным (и потому не накапливающимся и непосредственно не наблюдаемым), но необходимым промежуточным состоянием пути из А в В, — или, иными словами, что кинетика переходов А®В (и В®А) определяется высотой максимума свободной энергии на пути, соединяющем А и В.
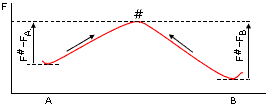
Рис.17-5. Преодоление свободно-энергетического барьера # при переходе из состояния "А" в состояние "В" (стрелка слева) и из "В" в "А" (стрелка справа). FA, FВ и F# — свободные энергии состояний А, В и "переходного" (барьерного, имеющего максимальную свободную энергию на пути процесса) состояния #.
При этом скорости перехода из А в В и из В в А суть
kА®В = k0 exp[-(F#-FA)/RT] kВ®А = k0 exp[-(F#-FВ)/RT]. | (17.1) |
Здесь FA, FВ и F# — свободные энергии состояний А, В и "переходного" # (Рис.17-5), а k0 — быстрота "элементарного шага" процесса, за которую часто принимают частоту теплового колебания RT/h (где h — постоянная Планка; при "обычных" температурах T~300oK, RT/h ~ 1013 сек-1), — с такой частотой, под воздействием тепловых колебаний, молекула предпринимает попытки преодолеть активационный барьер #.
Уравнения (17.1) применимы только к процессам, протекающим за время, много-много большее, чем время элементарного шага. И именно с такими процессами мы имеем дело при сворачивании белков.
Напомню, откуда следуют соотношения (17.1). Рассмотрим процесс А « # ® В. Так как "проток" молекул через барьер # идет медленно по сравнению со скоростью элементарного шага, то состояние # находится почти в термодинамическом равновесии с состоянием А. Иными словами, если в исходном состоянии А находится n молекул, то, по распределению Больцмана, в барьерном состоянии # находится n# = n exp[-(F#-FA)/RT] молекул. За время "элементарного шага" они скатятся с барьера # — половина в сторону А, половина в сторону В [а на их место придет примерно столько же (n#) молекул со стороны А — так что населенность # практически не изменится, будет квазистационарной]. Иначе говоря, для перехода половины всех n молекул через барьер в сторону В потребуется время, в (n/2)/(n#/2) = exp[+(F#-FA)/RT] раз превосходящее время элементарного шага. А поскольку время элементарного шага t0º1/k0, то характерная скорость реакции А « # ® В есть kА®В = k0 exp[-(F#-FA)/RT], а ее характерное время, — время, за которое около половины всех молекул перейдет из А в В, — есть
tА®В = 1/kА®В = (1/k0) exp[+(F#-FA)/RT]. | (17.2) |
Это же объяснение распространяется, естественно, и на kВ®А.
Важно, что температурная зависимость скорости реакции дает возможность судить об энергии переходного состояния. Для этого, согласно Аррениусу, вычисляют производную логарифма скорости реакции по обратной температуре (1/T):
d[ln(kА®В)] / d(1/T) = d[ln(k0) - (F#-FA)/RT]/(-T-2dT) ® - (E#-EA)/R . | (17.3) |
Здесь я воспользовался уже известной нам формулой d(F/T)/dT = - E/T 2 и пренебрег очень слабой зависимостью скорости элементарного шага от температуры. Последнее допустимо даже в простых химических реакциях, и тем более допустимо в белках, где энергии E# и EA — велики, так как определяются взаимодействиями множества частиц.
Рисунок 17-6 показывает изменение скорости сворачивания и разворачивания лизоцима при его тепловой ре - и денатурации.
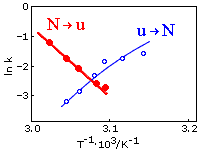
Рис.17-6. Аррениусовы графики для зависимости скорости тепловой денатурации и ренатурации лизоцима от обратной температуры (Т-1); графики взяты из статьи S. Segava & gihara, Biochemistry (1984) 23:2473-2488. Константы скорости (k) измеряются в сек-1. Ренатурация, скорость ku®N (эксперимент: точки о, и тонкая интерполяционная кривая); денатурация, скорость kN®u (эксперимент: точки ·, и жирная интерполяционная кривая). Середине плавления отвечает та температура, при которой ku®N = kN®u, т. е. где кривые пересекаются (около 1000/3.08 = 325оК). При меньших температурах (т. е. при больших Т-1 — справа от точки пересечения) превалирует сворачивание, при больших температурах (т. е. при меньших Т-1, слева от точки пересечения) — идет разворачивание.
Он показывает, что в самой середине перехода [там, где скорость сворачивания (ku®N) равна скорости разворачивания (kN®u), так что кривые для ln(ku®N) и ln(kN®u) пересекаются] — скорости обоих процессов близки к е-2.5 » 0.1 сек-1 (т. е. что здесь времена протекания и де-, и ренатурации — порядка 10 сек).
Кроме того, эта картинка показывает, что денатурация ускоряется по мере углубления в "область денатурации", а ренатурация ускоряется по мере углубления в "область ренатурации". И здесь содержится одна очень любопытная вещь.
Она состоит в том, что скорость тепловой денатурации kN®u падает с Т-1 (т. е. что она растет с температурой Т, что обычно для физико-химических реакций), а скорость сворачивания ku®N — наоборот, растет с Т-1 (т. е. она падает с температурой, что необычно для физико-химических реакций). По формуле (17.3) это означает, что E#-EN > 0, а E#-Eu < 0. Иначе говоря, Eu > E# > EN, т. е. энергия барьера, E#, лежит выше энергии нативного состояния, EN, но ниже энергии развернутого состояния, Eu (последнее необычно для химических реакций, где барьер по энергии выше чем и начальное, и конечное состояние реакции). Дальнейший анализ этого графика показывает, что такое же соотношение, Su > S# > SN, справедливо и для энтропий развернутого, переходного и нативного состояний при сворачивании белка. Это значит, что барьер между нативным и развернутым состояниями белка выглядит "обычным", энергетическим барьером, — если глядеть на него со стороны нативного состояния, и что он выглядит необычным, энтропийным барьером, — если глядеть на него со стороны развернутого состояния.
Впрочем, этого можно было ожидать, исходя из сделанного на прошлой лекции анализа свободно-энергетического барьера и причин фазового, типа "все-или-ничего" перехода между нативным и денатурированным состоянием белка (см. Рис.17-7, который воспроизводит уже известную вам схему изменения энергии, энтропии и свободной энергии по мере расширения глобулы).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
