
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Можно ли улучшить опознавание белковых структур? Да, если привлечь гомологи — т. е. предсказывать структуру белка уже не только по его аминокислотной последовательности, а по набору гомологичных последовательностей. Этот метод улучшил предсказания вторичных структур белков. Он широко и успешно применяется при предсказании вторичных структур РНК. При опознавании пространственных укладок белковых цепей его тоже начали применять — при этом отдается предпочтение той укладке, что более или менее хороша для всех гомологов. Правда, при этом мы получим не точную структуру данной цепи, а некоторую обобщенную, т. е. приближенную структуру, общую для всех гомологов.
Суммируем. Предсказание трехмерных укладок белковых цепей, с точки зрения физики, является поиском самой стабильной укладки рассматриваемой цепи (или, в современной реализации этого подхода, в "протягивании", threading'е — поиском структуры, наиболее совместимой с данной цепью). Такое предсказание возможно в принципе, и порой оно приводит к вполне удовлетворительному результату. Однако оно всегда имеет сложности, связанные с недостатком точной информации о репертуаре возможных укладок белковых цепей, о многих взаимодействиях в белковой цепи, с погрешностями в энергетических параметрах, и т. д.
В результате, однозначное и надежное предсказание укладки цепи по ее последовательности вряд ли возможно, — но зато можно выделить несколько наиболее вероятных мотивов укладки цепи и — что очень важно — отбраковать огромное множество других укладок. И такие предсказания могут быть уточнены с учетом дополнительной информации — в частности, о последовательностях цепей гомологичных белков и об их возможных укладках.
Мой сегодняшний рассказ касался в основном белков глобулярных. А что можно сказать о фибриллярных и мембранных?
В последовательностях фибриллярных белков — без всякого компьютера — просматриваются периодичности, характерные для их вторичных структур, — причем они настолько очевидны, что опознание вторичных структур этих белков обычно не составляет труда. А на этой основе удается предложить несколько возможных вариантов их упаковки в фибриллу.
Предсказание строения мембранных белков менее развито, чем предсказание строения белков глобулярных: слишком мало структур мембранных белков мы знаем. Опознать внутримембранные части таких белков довольно легко: это — сплошные (точнее — почти сплошные) гидрофобные блоки, длина которых диктуется толщиной мембраны. Однако принципы формирования укладки этих блоков в белке еще недостаточно выяснены (хотя известно главное; напомню: у мембранных белков ядро структуры — не гидрофобное, как у глобулярных, а полярное; гидрофобные же группы их контактируют с жиром мембраны). Поэтому работы по предсказанию пространственного строения мембранных белков только начинаются.
В заключительной части этой лекции я хочу кратко рассказать о белковой инженерии, точнее — о белковом дизайне, то есть о конструировании новых белков.
Олигонуклеотидный синтез и техника рекомбинантных ДНК дали возможность получения генов белков, не существовавших в природе; рентген и ЯМР позволили увидеть трехмерные структуры белков; а мощные ЭВМ и компьютерная графика позволили вступить в интерактивный диалог с этими пространственными структурами, — менять в них что-то и оценить последствия этих изменений. Объединившись, эти методы стали, соответственно, "руками", "глазами" и "мозгом" новой области молекулярной биологии — белковой инженерии. Ее стратегическая задача — создание знаний и методов, позволяющих получать белки с наперед заданной функцией и структурой. Трудно переоценить перспективность этих исследований для конструирования новых лекарств, катализаторов, для схемотехники и т. д.
Прицельный белково-инженерный эксперимент уже дал ответ на ряд фундаментальных вопросов. Показано, что за выбор пространственной структуры отвечают далеко не все детали плотной упаковки — структура белка выдерживает массу точечных мутаций; и не петли — если заменить или вырезать из белковой цепи участок, кодирующий петлю, то "рана" на теле глобулы обычно затягивается. О том, что целенаправленное введение точечных мутаций дает массу информации об энергетике белковой глобулы и промежуточных структур на пути ее самоорганизации, мы уже говорили; а о том, как они применяются для исследования активности белка — поговорим на следующей лекции.
Освоив введение мутаций в "природные" белки, белковая инженерия обратилась к дизайну, конструированию белковых молекул.
Задача дизайна — обратная по отношению к задаче предсказания структуры. Если при предсказании мы должны найти пространственную структуру, наиболее пригодную для рассматриваемой последовательности — то при дизайне мы должны найти, сконструировать последовательность, годную для создания желаемой пространственной структуры.
Вообще говоря, расчет искусственных конструкций может быть проще, чем предсказание структуры "натуральных" белков (расчет прочности "искусственной" башни проще, чем "натурального" дерева: она спроектирована так, чтоб поддаваться расчету!). А конструирование новых белков опирается на теорию белковых структур; их "строительными блоками" обычно служат последовательности, кодирующие мощные, внутренне стабильные и способные к эффективному слипанию a - и b-участки.
Белковый дизайн был поставлен на повестку дня в конце 70-х — начале 80-х годов, когда возникла техника создания искусственных генов и техника точного химического синтеза белковых цепей. В конце 80-х — начале 90-х годов при помощи прикидок, проб и ошибок были созданы простейшие белковые молекулы. Их архитектуры брались из природных белков, а их аминокислотные последовательности подбирались так, чтобы, не будучи гомологичными природным, стабилизировать эту архитектуру.
Первым был сделан — в группе ДеГрадо — четырехспиральный пучок (Рис.19-15). Дизайн проводился в тесном диалоге с экспериментом. Исследование окончательного варианта белка показало, что он спирален и глобулярен; и структура этого искусственного белка оказалась гораздо стабильнее, устойчивее к нагреву, чем структура любого "естественного" белка! Потом, правда, оказалось, что этот белок не плавился при нагревании потому, что был расплавленной глобулой с самого начала... Пришлось усилить твердую структуру этого искусственного белка, введя в него гистидины, связывающие ион. И вот этот-то белок-комплексон оказался уже твердым, как природные белки.
| Рис.19-15. Основные этапы дизайна четырехспирального пучка, сделанного в группе ДеГрадо. 1. Подбор коротких спиральных пептидов, способных слипаться в тетрамер. 2. Дизайн петель, сшивающих эти пептиды попарно, и отбор тех вариантов сшитых пептидов, что димеризуются. 3. Дизайн последней петли и отбор мономерного тетраспирального искусственного белка. Рисунок взят из [6]. |
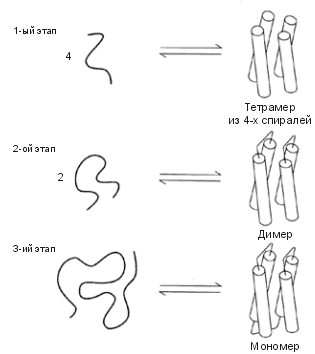
Долгое время все искусственные белки (кроме тех, что усиливались образованием ионных комплексов) представляли собой коллекцию отличных расплавленных глобул. Они обладали прекрасной вторичной структурой, они были очень компактны, — но им не хватало твердости.
Вопрос: Почему хотят сделать нормальный, "твердый" белок — а получается расплавленная глобула?
Видимо, потому, что все знают, как сделать стабильные вторичные структуры (лейцины и аланины, отороченные глутаминовыми кислотами с N-конца и лизинами с С-конца — получится a-спираль; побольше валинов, изолейцинов и треонинов — получится b-структура); и все знают, как заставить эти a - и b-структуры слипаться: надо сделать на них сплошные поверхности из гидрофобных групп (см. Рис.19-6); и как заставить эти белки не агрегировать: надо сделать противоположные поверхности этих a - и b-структур из гидрофильных групп. Но вот для чего рецептов пока никто не сделал — это для создания плотной упаковки боковых групп в гидрофобном ядре белка. Вот эта-то упаковка и не получается. А без нее — при наличии слипающихся, но не плотно слипающихся вторичных структур, — получается лишь расплавленная глобула...
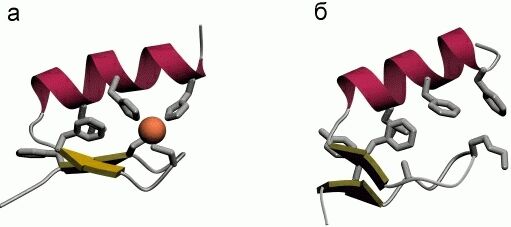
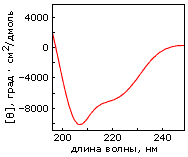
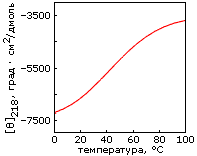
Однако, по-видимому, задача дизайна плотной упаковки может быть решена. Имея хороший алгоритм компьютерного перебора астрономического числа возможных упаковок боковых групп и отсечения бесперспективных вариантов, расчет плотной упаковки — для маленького белка — может быть проведен. По крайней мере, Дахийат и Майо его провели и создали — в 97 г. — твердый (без всякого дополнительного комплексообразования с ионом) маленький белок (Рис.19-16). Этот белок был создан на основе архитектуры "цинкового пальца" (широко распространенного ДНК-связывающего мотива), — но без иона Zn, держащего архитектуру природного цинкового пальца. При этом искусственный белок FSD-1 Дахийата и Майо имеет очень невысокую (20%) гомологию с природным цинковым пальцем, — и лишен цинк-связывающего центра. И тем не менее он тверд при низких температурах (его структура исследована при помощи ЯМР). Однако он плавится в довольно широком температурном диапазоне, т. е. заметно менее кооперативно (Рис.19-16г), чем аналогичные ему природные белки.
в г

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
