
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
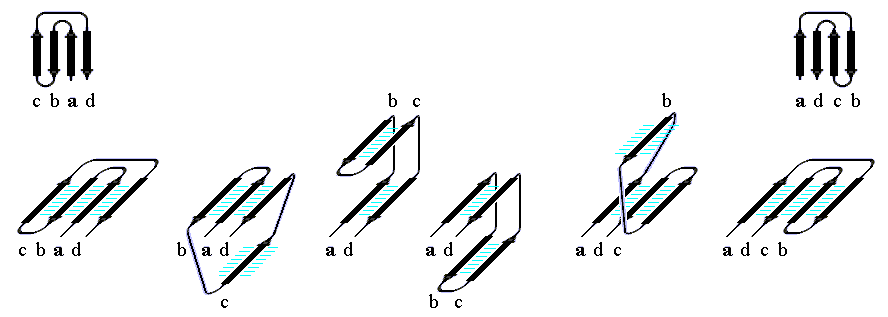
Рис.13-15. Различные пространственные "abcd" структуры Ефимова с топологией греческого ключа. Обратите внимание на правый ход суперспирали, состоящей из двух параллельных b-тяжей одного b-листа и лежащего между ними в цепи b-тяжа другого листа (т. е. на суперспираль из b-тяжей b-c-d на втором слева внизу и на суперспираль из тяжей a-b-c на второй справа внизу схемах). Именно такое — "правовинтовое" — соединение параллельных b-тяжей одного b-листа типично для белков; обратное, "левовинтовое" встречается очень редко.
Все такие характерные, часто встречающиеся в белках структуры (шпильки, меандр, греческий ключ, abcd структуры и т. д.), сложенные из соседних по цепи элементов b (и/или a) структуры, часто называют "супервторичными" структурами.
Лекция 14
Перейдем теперь к a-белкам — белкам, сложенным из a-спиралей. Их труднее классифицировать, чем b-белки. Дело в том, что взаимное расположение b-участков в листах поддерживается водородными связями в главной цепи (а она — повсюду одинакова), в то время как расположение a-спиралей в глобуле поддерживается плотной упаковкой их боковых групп, которые весьма разнообразны по размеру. Поэтому a-спирали не уложены в такие ровные, стандартные листы, как b-тяжи.
Проще других устроены те a-белки, где a-спирали длинны. Такие спирали образуют пучки: они лежат (почти) параллельно или антипараллельно (в общем, "колинеарно") друг к другу. С пучками спиралей мы уже встречались в фибриллярных и мембранных белках.

Рис.14-1. Три сходных по архитектуре ("четырехспиральный пучок"), но разных по функции a-спиральных белка: цитохром c', миогемэритрин и белок оболочки вируса табачной мозаики. Показана как белковая цепь, так и кофакторы: скелетные модели — гем (в цитохроме) и фрагмент РНК (в белке оболочки вируса), оранжевые шарики — ионы железа (в геме цитохрома и в миогемэритрине), красный шарик — связанный железом кислород (в миогемэритрине). Общая архитектура таких "пучков" напоминают коллинеарную упаковку b-листов. На топологической схеме (внизу) дан вид на белок "снизу" (с торцов структурных сегментов). Спирали занумерованы цифрами. Кружки - торцы a-спиралей. Крестик соответствует N-концу сегмента (т. е. он "идет от нас"), точка — его С-концу (т. е. он "идет к нам"). Ход петель, соединяющих структурные сегменты, показан черной линией, если петля обращена к нам, и светлой, если она находится на противоположной стороне укладки. Цифры на схеме указывают порядок структурных сегментов в цепи.
На Рис.14-1 показаны три четырехспиральных a-белка. Эти, столь похожие между собой белки исполняют разные функции: один из них, цитохром, связывает электрон; другой (миогемэритрин) связывает кислород, а третий (белок облочки вируса табачной мозаики) связывает гораздо более крупные молекулы — другие оболочечные белки и РНК. В первых двух случаях какую-то общность действия проследить можно, так как оба белка что-то переносят в дыхательной цепи. Правда, сходство здесь весьма отдаленное: в цитохроме полипептид — а именно о его укладке идет речь — связывает гем, тот — ион железа, а уж оно-то и хватает электрон; а в миогемэритрине полипептид связывает железо непосредственно, без гема, — и уже два иона железа, а не один, — которые и хватают кислород. Итак, у миогемэритрина и цитохрома некую общность функции, пусть с большой натяжкой, но можно найти, — но, конечно, у их действия нет ничего общего с действием РНК-связывающего белка оболочки вируса, — несмотря на очень сходную архитектуру структурного каркаса, четырехспирального пучка. Причем сходство распостраняется не только на архитектуру — четырехспиральный пучок — но и на ход цепи сквозь эту архитектуру, т. е. на мотив укладки цепи. Последнее подчеркивается общей для всех трех белков топологической схемой, приведенной внизу рисунка (схема изображает вид пучка с торца).
Итак, при одинаковом мотиве укладки цепи белки могут работать совсем по-разному. А вот миогемэритрин и классический кислород-связывающий белок миоглобин (Рис.14-2) делают одно и то же дело (только первый — в червях, а второй — в хордовых), — но их архитектуры совсем непохожи, — за исключением того, что оба они — a-белки. Но в миогемэритрине все a-спирали лежат параллельно, а в миоглобине спирали организованы в два перпендикулярных слоя.
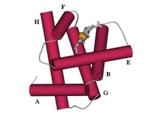
Рис.14-2. Структура глобина: скрещенные слои по три a-спирали в каждом. Спирали A, E и F (они занумерованы буквами по порядку нахождения в цепи) находятся в верхнем слое, спирали H, G и B — в нижнем. Короткие (из 1 — 2 витков каждая) спирали С и D не изображены, так как они не консервативны в глобинах. В щели верхнего слоя находится гем. Такие "скрещенные слои" напоминают ортогональную упаковку b-листов.
Это — еще один пример того, что сходная функция может осуществляться белками с разной архитектурой, а белки одинакового устройства могут заниматься совсем разным делом.
Я, как и раньше, хочу привлечь ваше внимание к нетривиальным случаям малой связанности структуры и функции белка, понимая, что вы и так знаете, что родственные белки — например, миоглобин и другие глобины — похожи по строению и занимаются одним и тем же делом.
Сравнивая миоглобин с миогемэритрином, я хочу обратить ваше внимание еще на то, что активный центр обоих этих белков (в первом случае этот центр — гем с ионом железом внутри, во втором — два иона железа), — этот центр локализован в "архитектурном дефекте" структуры — в данном случае, в щели между раздвинутыми спиралями.
Рассмотренные выше "пучки" типичны для упаковки весьма длинных a-спиралей. Они встречаются как в водорастворимых глобулярных белках, так и в фибриллярных и в мембранных белках. При этом ядро белка, заключенное между a-спиралями (гидрофобное — в водорастворимых глобулярных и фибриллярных белках, гидрофильное — в мембранных) имеет вытянутую, квазицилиндрическую форму.
Однако для относительно коротких спиралей, типичных для глобулярных белков — их длина обычно составляет около 20![]() при диаметре 10
при диаметре 10![]() — более характерна квазисферическая укладка спиралей вокруг "квазишарового" гидрофобного ядра, сложенного из сидящих на этих спиралях боковых групп..
— более характерна квазисферическая укладка спиралей вокруг "квазишарового" гидрофобного ядра, сложенного из сидящих на этих спиралях боковых групп..
Типичная упаковка спиралей в глобулярном белке показана на Рис.14-3. Ее нельзя описать в терминах параллельной и перпендикулярной упаковки спиралей — большинство межспиральных контактов имеет угол около 40 — 600.
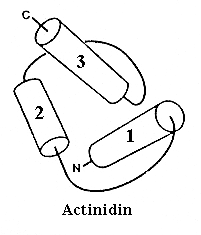
Рис.14-3. Типичная упаковка спиралей в глобулярном белке: N-концевой домен актинидина (ход петель прослежен очень грубо). Обратите внимание, что архитектуру этого домена невозможно описать в терминах колинеарных и ортогональных упаковок a-спиралей.
Однако и такие сложные укладки можно неплохо описывать и классифицировать при помощи модели "квазисферических многогранников", которую мы с Алексеем Мурзиным предложили 10 лет назад. Для примера, — покажу (Рис.14-4), как эта модель описывает только что показанную (на Рис.14-3) a-спиральную глобулу. Эти два последних рисунка, кстати, взяты из опубликованной в "Nature" рецензии на нашу работу.
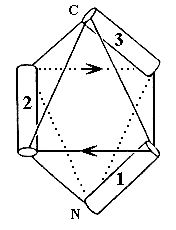
Рис.14-4. Размещение a-спиралей на ребрах квазисферического многогранника, моделирующее N-концевой домен актинидина. Стрелки указывают ход соединяющих a-спирали петель.
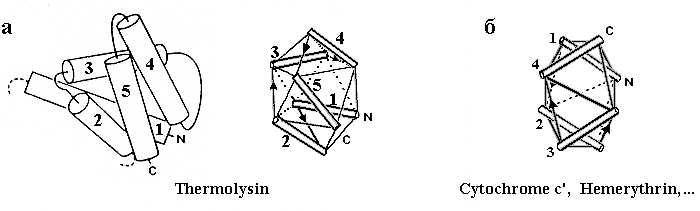
Рис.14-5. Другие примеры того, как упаковка спиралей в глобулярном белке описывается моделью квазисферических многогранников. (а) С-концевой домен термолизина и его модель, основанная на размещении спиралей на ребрах квазисферического многогранника; (б) модель четырехспиральной глобулы, показанной на Рис.14-1.
Не откажу себе в удовольствии показать и еще пару рисунков из той же рецензии (Рис.14-5а).
Кстати, модель квазисферических многогранников довольно прилично описывает и довольно длинные спиральные пучки — те, что мы видели на Рис.14-1. Это демонстрируется рисунком 14-5б.
Суть модели квазисферических многогранников заключается в том, что она концентрирует наше внимание на расположении спиралей вокруг шарового ядра глобулы. Модель учитывает только, что a спирали — твердые вытянутые частицы — плотно окружают ядро; и что полярные концы спиралей должны находиться на поверхности глобулы. Каждая упаковка спиралей может моделироваться многогранником (Рис.14-6), каждая вершина которого соответствует как бы половине спирали. Самые компактные, "квазисферические" многогранники (Рис.14-7) описывают компактные глобулы. Упаковки, близкие к идеальным, и наблюдаются в глобулярных a-белках. При этом каждому данному числу спиралей отвечает один многогранник, а в его рамках существует несколько (от двух до десяти) типов укладок, соответствующих различным размещениям осей спиралей на ребрах этого многогранника. Среди этих укладок есть и рассмотренные выше "пучки спиралей", и "скрещенные слои".

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
