
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Продолжим качественное обсуждение изменения скорости сворачивания с повышением стабильности глобулярной структуры. Предположим, что изменение внешних условий (температуры или состава растворителя) повышает стабильность глобул — всех глобул, "правильно" и "неправильно" свернутых, а на только нативной — по сравнению с развернутым состоянием цепи.
Если первоначальный рост стабильности глобул относительно клубка ускоряет сворачивание самой стабильной структуры, так как ее стабильность растет, а ее конкуренты все еще остаются нестабильными относительно исходного, развернутого состояния цепи, — то дальнейший рост стабильности глобул делает уже не одну только самую стабильную из них, но и многие из компактных, но "неправильно свернутых" форм более стабильными, чем развернутое состояние цепи (Рис.18-5б): появляются метастабильные "ловушки", способные удержать цепь и замедлить ее попадание в нативную структуру. При этом барьеры на путях, ведущих к некоторым "ловушкам", могут быть ниже, чем на путях, ведущих к самой стабильной структуре; тогда возможно быстрое скатывание к метастабильной структуре (или структурам) глобулы — и очень медленный переход к истинно стабильной структуре.
Стоит отметить, что метастабильные "ловушки" перехватывают сворачивание к стабильной структуре тем эффективнее, чем меньше энергетическая щель между нативной структурой и прочими (метастабильными) укладками цепи. Это еще раз подчеркивает, что быстро находить свою стабильную структуру может только такая цепь, у которой стабильная пространственная структура возникает путем фазового перехода первого рода, — т. е. цепь, у которой энергия ее самой стабильной укладки отделена достаточно большой щелью от энергий конкурирующих с ней структур.
Хочу отметить, что сходство термодинамики самоорганизации белков с фазовыми переходами первого рода экспериментально установлено в работах , а сходство кинетических аспектов этих явлений — в компьютерных экспериментах и .
Здесь у вас естественно возникает вопрос: — Что будет, если у цепи не одна структура отделена от прочих большой энергетической щелью, — а, скажем, две?
Ответ: — Если обе эти структуры стабильны по сравнению с клубком — первой из них свернется та, к которой идет лучший (с немного более низким барьером) путь сворачивания. Однако, если эта структура хоть немного менее стабильна, чем другая, — последует очень медленный (так как по пути придется разворачивать метастабильную структуру, см. Рис.18-5б) переход в наиболее стабильную форму. Этот переход похож на полиморфные переходы в кристаллах (вспомним "оловянную болезнь" — переход белого олова в серое: эта "болезнь" порой уничтожала, при наступлении холодов, целые склады оловянных пуговиц). По-видимому, таких "полиморфных" белков должно быть мало (теоретические оценки показывают, что аминокислотные последовательности, кодирующие одну выделенную стабильную укладку цепи, — редки, а кодирующие сразу две такие укладки — редки в квадрате).
Однако есть основания полагать, что некоторые белки — серпины (есть такие ингибиторы сериновых протеаз) и, возможно, прионы (они вызывают "бешенство коров") — ведут себя именно так. В последнем случае полиморфизм осложнен агрегацией (слипанием), — агрегацией, которой способствует — или которой наводится — "новая", b-структурная форма белка. При агрегации растет эффективная длина цепи фолдона (N), что (согласно сделанным выше оценкам) делает возникновение новой фазы очень медленным. Но зато раз возникший зародыш новой формы агрегированного белка втягивает в агрегат и переводит в эту новую форму все остальные белковые молекулы (по существу — так же, как при "оловянной чуме"). Эта-то агрегация прионов в клетках мозга и приводит в конце концов к летальному исходу при "коровьем бешенстве", почесухе и родственных им болезням с огромным инкубационным периодом. Говорят, к таким болезням относится и старческий маразм...
Впрочем, есть и более простой, "мирный" и хорошо изученный лабораторный пример того же рода: водорастворимый аминокислотный полимер поли(лизин) в пробирке, при рН<10 и температуре 20-50ОС, быстро (за миллисекунды) переходит из клубка в a-спираль, а потом — гораздо медленнее — из a-спирали в b-структуру. Последний переход сопровождается агрегацией и может занимать часы, недели и более, — его время экспоненциально растет с падением стабильности b-структуры; об этом мы уже говорили, обсуждая кинетику образования b—листов.
В заключение обратимся еще раз к энергетическому ландшафту белковой цепи (Рис.18-1) и посмотрим, как самая стабильная структура автоматически пролагает к себе быстрые, преодолевающие парадокс Левинталя пути сворачивания.
Через очень холмистый (скорее — даже скалистый) ландшафт (Рис.18-1) от каждого энергетического минимума широко расходится "воронка" (или, точнее, сеть) более или менее гладких "дорожек", соответствующих разным путям последовательного (Рис.18-2) сворачивания структуры, соответствующей рассматриваемому минимуму. На всех этих путях не происходит перестройки уже свернувшейся части глобулы, и потому на них нет больших ухабов (а маленькие — не помеха, если температура не слишком низка). Двигаясь по этим дорожкам к энергетическому минимуму, молекула выигрывает энергию, но, одновременно, фиксируя свою укладку, — теряет энтропию (Рис.18-3). Чем глубже энергетический минимум — тем круче ведущие к нему дорожки, тем легче преодолевать, идя по ним, "энтропийное сопротивление", возникающее при фиксации укладки скатывающейся в энергетический минимум цепи. Сила этого сопротивления — поскольку оно энтропийное — пропорциональна температуре. При очень высокой температуре оно выталкивает цепь из всех энергетических минимумов, так что цепь не сворачивается вообще. При очень низкой температуре энтропийное сопротивление мало — цепь скатывается в любой ближайший минимум, долго из него выбирается, снова куда-то скатывается, — и никак не может дойти до глобального энергетического минимума (Рис.18-5б). И, наконец, при оптимальной для сворачивания температуре — энтропийное сопротивление преодолевается на путях только к одному, самому глубокому минимуму — куда цепь и попадает достаточно быстро (Рис.18-5а).
Все изложенные соображения о компенсации энергией энтропии по пути самоорганизации и о том, что это решает парадокс Левинталя, применимы и к образованию нативной структуры белка из расплавленной глобулы. Но там сделать какие-либо количественные оценки гораздо сложнее. К тому же хочу подчеркнуть, что эксперимент не показывает принципиального, многопорядкового ускорения самоорганизации белка при образовании расплавленных глобул. Поэтому я и ограничился только более простым случаем образования нативной структуры прямо из клубка.
Лекция 19
С тех пор как стало понятно, что аминокислотная последовательность белковой цепи определяет ее пространственную структуру — возникла проблема предсказания этой структуры по последовательности аминокислотных остатков в белковой цепи.
Чем вызвана потребность в предсказании белковых структур, — кроме чисто интеллектуального интереса: удастся это сделать или нет? Тем, что экспериментально пространственную структуру белка определить куда труднее, чем его аминокислотную последовательность. А понимание механизма действия белка, подбор искусственных ингибиторов или активаторов к нему, — и часто даже просто определение того, чем он занимается в клетке, — настоятельно требует знания его пространственной структуры...
И — конечно же! — интерес к предсказанию пространственных структур белков подогревается воспоминанием о том, какое решающее значение имело предсказание строения двойной спирали ДНК для понимания всего генетического механизма.
Сейчас известно уже порядка сотни тысяч белковых последовательностей. Но "всего" для нескольких тысяч из них, т. е. всего для нескольких процентов определены, рентгеном или ЯМР, их пространственные структуры. При этом многие из недавно определенных последовательностей просто считаны с ДНК или РНК, т. е. никто не определил на опыте, чем занимаются сделанные из них белки.
Что же можно сказать о трехмерных структурах тех последовательностей (я их буду называть "новыми"), для которых эксперимент — рентген или ЯМР — еще не сказал своего слова?
Прежде всего возникает мысль о предсказании трехмерной структуры "новой" последовательности на основании родственного сходства — или, как говорят, "гомологии" ее первичной структуры с какими-то из "старых" последовательностей, пространственное строение коих уже расшифровано. Опыт показывает, что даже не очень сильного сходства последовательностей достаточно для очень хорошего сходства пространственных структур: как говорят, пространственная структура более консервативна, чем аминокислотная последовательность.
Установление гомологии первичных структур — действительно, очень мощный метод выяснения родства структур (причем не только белков, а и фрагментов ДНК и РНК — но я буду говорить о белках).
Однако надежно он работает, надежно устанавливает сходство первичных пространственных структур только на достаточно близких последовательностях. Этот случай иллюстрируется Рис.19-1.
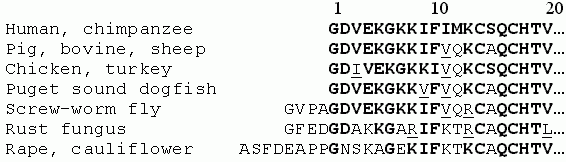
Рис.19-1. Гомологичные аминокислотные последовательности N-концевых фрагментов цитохромов c различных митохондрий и хлоропластов эукариотов. Жирным шрифтом выделены остатки, идентичные оным в человечьем (human) белке, подчеркнуты — сходные с ними. Выравнивание аминокислотных последовательностей взято из [6].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
