
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Биосинтез коллагена, его последующая модификация и образование зрелой структуры коллагеновой фибриллы хорошо изучены (Рис.11-8). Замечу, что сам по себе коллаген не способен к спонтанной и при этом правильной самоорганизации своей пространственной структуры in vitro — так же, как и фиброин шелка, — и в отличие от глобулярных белков, о которых я буду говорить потом. Для самоорганизации нужен проколлаген, включающий, кроме коллагеновых нитей, глобулярные головки и хвостики. Лишенные головок и хвостиков коллагеновые нити, самоорганизуясь из развернутого состояния in vitro, тоже складываются в тройные спирали — но "неправильные", без присущей нативному коллагену гетерогенности тройной спирали (включающей нити двух сортов), без присущего ему регистра (т. е. с неправильным сдвигом нитей относительно друг друга), и т. д.
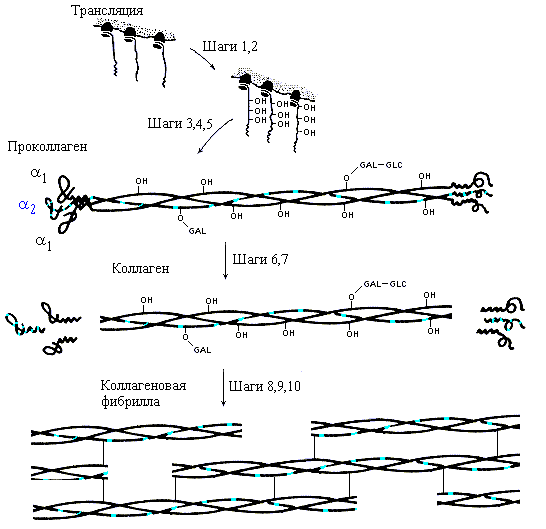
Рис.11-8. Образование коллагена in vivo. Шаг 1. Биосинтез про-a1-цепей и про-a2-цепей (по 1300 остатков в каждой) в пропорции 2:1. Шаг 2. Гидроксилирование некоторых остатков Pro и Lys. Шаг 3. Присоединение сахаров (GLC-GAL) к гидроксилированным остаткам. Шаг 4. Образование тримера и SS связей на его концах. Шаг 5. Образование тройной спирали в середине проколлагена. Шаг 6. Секреция проколлагена во внеклеточное пространство. Шаг 7. Отщепление глобулярных частей. Шаги 8-10. Спонтанное образование фибрилл из тройных суперспиралей, окончательная модификация аминокислотных остатков и образование ковалентных сшивок модифицированных остатков коллагеновых цепей. Картинка взята из [3] и адаптирована.
В заключение, я хочу подчеркнуть, что фибриллярные белки устроены относительно просто в силу периодичности своей первичной и, в силу этого, — также и своей вторичной структуры.
12
Рассмотрим теперь мембранные белки. Они тоже — я имею в виду их трансмембранные части — устроены довольно просто, почти как фибриллярные белки.
Мембраны создают поверхность клетки, а внутри нее, — разные замкнутые объемы — или, как говорят, компартменты. Мембраны состоят из жира (липидов) и белков (Рис.12-1). Особая роль мембранных белков (а они составляют до половины веса мембраны) — обеспечивать транспорт через нее различных веществ, а также сигналов. Мембрана — это, так сказать, "изолятор", а белки — точнее, как мы увидим, каналы в них — "проводники". Эти проводники специфичны, каждый из них пропускает через мембрану только определенные молекулы (или — по-видимому, посредством небольшого изменения конформации белка — сигналы от определенных молекул).

Рис.12-1. Белки в мембране. Серым цветом выделены внемембранные домены. Внутримембранные части белка практически не содержат нерегулярных участков цепи.
Истинно мембранные белки "живут" внутри мембраны — там воды практически нет, так что внутримембранные части таких белков должны состоять (и состоят, как мы увидим) из регулярной вторичной структуры, причем размер этих частей ограничен толщиной мембраны.
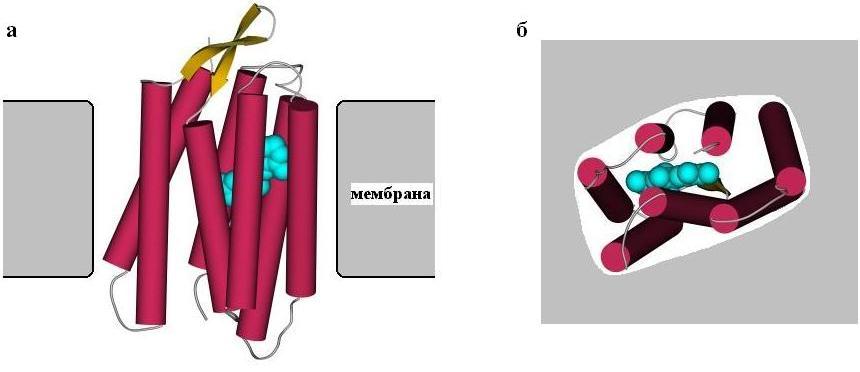
Рис.12-2. Бактериородопсин в мембране (а - вид вдоль мембраны, б - вид на мембрану сверху). Цилиндрами показаны семь спиралей этого белка. Показаны и соединяющие эти спирали петли, а также (голубым цветом) молекула ретинола, прикрепленная внутри бактериородопсина.
Посмотрим, на нескольких примерах, как устроены мембранные белки. Вообще надо сказать, что мы еще знаем мало структур мембранных белков, — их расшифровано меньше десятка, — так как эти белки плохо растворимы в воде (приходится использовать детергенты и т. д.), и их очень трудно кристаллизовать, так как они склонны слипаться неупорядоченно.
Структура бактериородопсина — он проводит протон через мембрану — показана на Рис.12-2; эта структура построена на основе анализа множества электронных микрофотографий очень высокого разрешения, так как получить трехмерные кристаллы бактериородопсина не удалось.
Мы видим, что трансмембранная часть бактериородопсина сложена из семи регулярных a-спиралей, идущих от одного до другого края мембраны и образующих слегка наклоненный к ее плоскости пучок, а одинокая b-шпилька и все нерегулярные участки цепи (соединяющие спирали петли) выходят из мембраны.
Высокая регулярность укладки остова трансмембранной белковой цепи закономерна: в "жирном", почти безводном липидном окружении цена каждой водородной связи очень высока, что заставляет белковую цепь, уж если она попадает в мембрану, принимать структуры с полностью завязанными водородными связями — т. е. либо a-спираль, либо b-структуру (точнее, b-цилиндр, не оставляющий края b-листа открытыми — см. ниже).
Сидящие на a-спиралях бактериородопсина гидрофобные группы обращены "наружу", к (тоже гидрофобным) липидам мембраны. Полярные же группы — их немного — обращены внутрь узкого канала, по которому идет протон. Протонная проводимость осуществляется при содействии прикрепленной внутри пучка спиралей молекулы кофактора — ретинола.
Аналогичные, устроенные как полый пучок спиралей поры могут образовываться в других случаях и из отдельных a-спиральных трансмембранных пептидов.
Посмотрим еще на один трансмембранный белок, порин (Рис.12-3). Он также высоко регулярен и имеет вид широкого цилиндра, сложенного из b-структуры. Обратите внимание, что b-лист здесь образует замкнутый b-цилиндр, что позволяет избежать "незавязанных" водородных связей, типичных для краев плоского b-листа. В этом цилиндре 16 очень длинных b-участков, а диаметр проходящей по его центру довольно широкой поры — около 15![]() . В сторону поры обращены боковые группы полярных остатков, входящих в b-тяжи, а чередующиеся с ними в b-тяжах неполярные остатки обращены своими боковыми группами в мембрану.
. В сторону поры обращены боковые группы полярных остатков, входящих в b-тяжи, а чередующиеся с ними в b-тяжах неполярные остатки обращены своими боковыми группами в мембрану.
Порин служит проводником полярных молекул — кстати, не очень селективным.
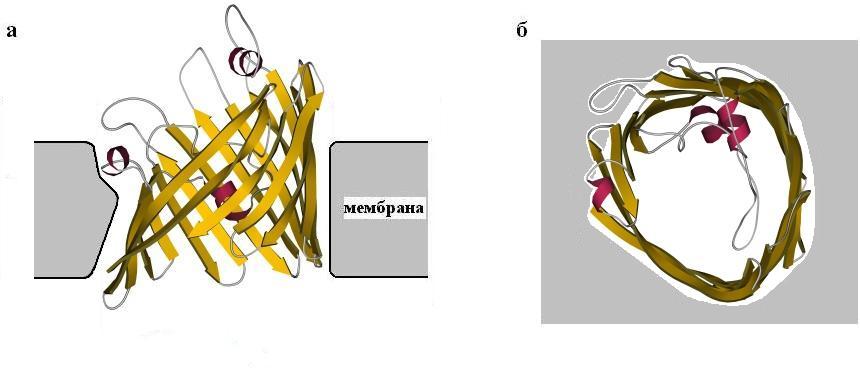
Рис.12-3. Порин. (а) Вид вдоль плоскости мембраны (расположение липидов в ней показано очень схематично). (б) Вид поперек плоскости мембраны.
Селективность проводимости, т. е. специфичность действия мембранных белков во многом связана с тем, что полярные, а тем более заряженные группы "сами по себе" проникают внутрь мембраны с большим трудом.
Вы помните, что свободная энергия заряда q в среде с диэлектрической постоянной e равна +q2/2er, где r — Вандерваальсов радиус заряда. Легко прикинуть, что при q, равном электронному заряду, и r =1.5 ![]() (характерный радиус однозарядного иона), величина +q2 /2er близка к 1.5 ккал/моль при e=80 (т. е. в воде), а при eмембр=3 (т. е. внутри "чистой", сложенной лишь из липида мембраны), величина +q2/2er близка уже к 37 ккал/моль. Итого — повышение свободной энергии на DF = +35 ккал/моль. Вероятность набрать такую свободную энергию есть, по формуле Больцмана, exp(-DF/kT) = exp(-35/0.6) = 10-25. Значит, успешной будет лишь одна из 1025 атак мембраны ионом. А так как "атака" длится не менее 10-13 сек (это — время теплового колебания, как мы помним), то для проникновения иона через чисто липидную мембрану нужно по крайней мере порядка 1012 секунд, или порядка десятка тысяч лет... Все это делает чисто липидную мембрану практически непроницаемой для ионов.
(характерный радиус однозарядного иона), величина +q2 /2er близка к 1.5 ккал/моль при e=80 (т. е. в воде), а при eмембр=3 (т. е. внутри "чистой", сложенной лишь из липида мембраны), величина +q2/2er близка уже к 37 ккал/моль. Итого — повышение свободной энергии на DF = +35 ккал/моль. Вероятность набрать такую свободную энергию есть, по формуле Больцмана, exp(-DF/kT) = exp(-35/0.6) = 10-25. Значит, успешной будет лишь одна из 1025 атак мембраны ионом. А так как "атака" длится не менее 10-13 сек (это — время теплового колебания, как мы помним), то для проникновения иона через чисто липидную мембрану нужно по крайней мере порядка 1012 секунд, или порядка десятка тысяч лет... Все это делает чисто липидную мембрану практически непроницаемой для ионов.
Другое дело, если в мембрану встроен белок, а внутри него образован более или менее широкий, заполненный водой канал, по которому ион может идти, не покидая среду с высокой диэлектрической проницаемостью. Правда, и в канале ион будет чувствовать низкую диэлектрическую проницаемость окружающей канал мембраны, но эффект будет уже много меньше — грубая оценка показывает, что он порядка +q2/(4eмембрR), где R — радиус канала, а eмембр =3. Легко прикинуть, что при этом канал радиусом »2 ![]() в мембране преодолевается ионом за секунду, а »3
в мембране преодолевается ионом за секунду, а »3 ![]() — за долю миллисекунды.
— за долю миллисекунды.
Пропускание иона через канал регулируется наличием в нем центров, способных притянуть данный ион и тем уменьшить барьер, который ему приходится преодолевать.
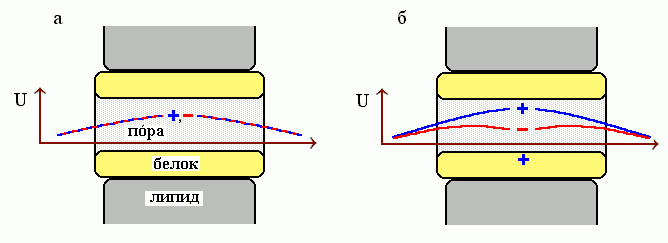
Рис.12-4. Схема трансмембранной поры (мембрана в данном случае расположена вертикально а пора — горизонтально), и электростатическая свободная энергия U для положительно ( .............. + .............. ) и отрицательно ( .............. - ..............) заряженных ионов. (а) На внутренней поверхности поры нет заряда. (б) На внутренней поверхности поры — положительный заряд.
Например, наличие на белке вблизи канала хоть какого-то положительного заряда ускоряет пропускание отрицательно заряженных ионов и резко замедляет пропускание ионов, заряженных положительно (Рис.12-4), а наличие отрицательного заряда — наоборот, ускоряет пропускание положительных ионов и замедляет пропускание отрицательных. Этот эффект служит основой селективности той проводимости, что обеспечивается мембранными белками.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
