
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
И "упаковки", и "мотивы укладки" цепи в белковую глобулу сосредотачивают наше внимание не просто на всевозможных (рыхлых, ажурных и т. п.) комплексах из структурных сегментов, а лишь на тех, где они, эти структурные сегменты, уложены плотно. Так мы очерчиваем те области конфигурационного пространства, что соответствуют плотной (но без стерического перекрывания) упаковке белковой цепи в глобулу — т. е. окрестности достаточно глубоких минимумов энергии невалентных взаимодействий. Они дают возможность не только классифицировать уже известные белковые структуры, но и предвидеть новые, еще не найденные в природе.
Здесь уместно сделать пояснение. Когда я буду говорить о классификации белковых структур, об их сходстве и т. д. — я буду иметь в виду не тривиальные вещи типа того, что все глобины похожи друг на друга, независимо от того, работают ли они в человеке или в морском червяке. Это, конечно, так, и белки можно классифицировать по филогенетическим семействам, внутри которых функции и, главное, аминокислотные последовательности белков варьируют не слишком сильно. Однако часто сходными пространственными структурами обладают белки, эволюционно никак, по всем тестам, не связанные между собой. И вот это, чисто структурное сходство я буду подчеркивать.
Начнем с изучения строения b-белков. b-структурные домены устроены, пожалуй, проще других: вытянутые участки цепи собраны обычно в два, реже — в несколько b-листов, уложенных друг на друга. Иными словами, "штабеля" вторичной структуры в b-белках выглядят довольно просто.
В b-белках преобладает антипараллельная b-структура.
Так как белки собраны из асимметричных (L) аминокислот, то вытянутые участки слегка скручены сами по себе. Причем, так как минимум энергии вытянутой конформации лежит, как вы помните, на карте Рамачандрана выше диагонали, — вытянутые участки имеют левовинтовую скрученность. В итоге объединения, при помощи водородных связей, скрученных b-участков в листы, — определенным образом скручены и эти b-листы (Рис.13-5). Их поверхность напоминает пропеллер. Угол между смежными вытянутыми участками b-листа составляет около -250. Значит, этот пропеллер выглядит как левовинтовой, если смотреть на него поперек хода b-участков (см. Рис.13-5а). И то же скручивание является правовинтовым, если смотреть вдоль хода b-участков (см. Рис.13-5б). Так обычно и делают, — смотрят вдоль хода b-тяжей и говорят, что у b-листа - правопропеллерная скрученность.
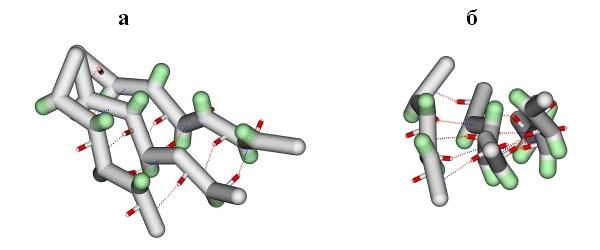
Рис.13-5. Лист b-структуры, вид поперек (а) и вдоль (б) b-тяжей. Лист имеет складчатую поверхность (она подчеркивается выступающими Сb-атомами, окрашенными в зеленый цвет), и — обычно — правопропеллерную (если смотреть вдоль тяжей) скрученность. Водородные связи между b-тяжами изображены пунктиром.
Существует два основных типа упаковки b-листов: продольная и ортогональная (Рис.13-6а и 13-6б). В обоих случаях b-листы упакованы "лицом к лицу", и гидрофобное ядро домена заключено между ними, но в первом случае листы повернуты друг относительно друга слабо — на угол около -300 (плюс-минус 10-150), а во втором — сильно, на 900 (плюс-минус 10-150); углы, лежащие вне этих двух диапазонов (в частности, углы около +300), наблюдаются редко.
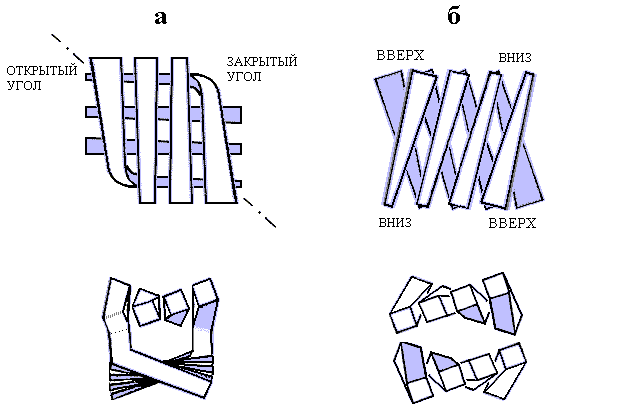
Рис.13-6. Ортогональная (а) и продольная (б) упаковка b-листов. Вид сверху и с торца. На виде сверху — приближение b-тяжа к читателю показано его (тяжа) расширением. Штрихпунктир — ось ортогонального b-бочонка. На оси находятся оба "открытых" угла этой упаковки. Здесь b-листы наиболее раздвинуты. В двух других, "закрытых" углах листы наиболее сближены; здесь цепь изгибается и переходит из слоя в слой. При ортогональной упаковке гидрофобное ядро имеет приблизительно цилиндрическую форму. В продольной упаковке, наоборот, ядро плоское, расстояние между скрученными листами меняется мало, а поворот одного листа относительно другого позволяет гидрофобным поверхностям лежащих друг над другом скрученных b-тяжей соприкасаться на большой длине. Картинка, с небольшими изменениями, взята из C. Chothia & A. V.Finkelstein, Annu. Rev. Biochem. (1990) 59:1007-1039.
При ортогональной упаковке (Рис.13-6а) b-участки скручены и обычно несколько изогнуты, так что общая форма "штабеля" начинает напоминать цилиндр с сильным наклоном b-участков к оси. Такую упаковку часто так и называют, — b-цилиндром или b-бочонком, хотя в b-цилиндрах, сложенных из антипараллельной b-структуры (в отличие от b-цилиндров, сложенных из параллельной b-структуры, о которых речь впереди), — в данном случае сеть водородных связей часто разорвана (или надорвана) на противоположных боках бочонка, что позволяет довольно четко выделить два b-листа. В "закрытом" углу такой упаковки b-участки обоих листов близко сходятся, так что цепь перетекает из листа в лист ценой изгиба и поворота на 900: можно сказать, что единый b-лист согнут и уложен сам на себя. В противоположных — "открытых" — углах b-листы расходятся, а образующаяся щель обычно заполняется или a-спиралью, или нерегулярными петлями, — или активным центром, как в ретинол-связывающем белке (Рис.13-7).

Рис.13-7. Ретинол-связывающий белок: пример ортогонально упакованного b-листа. Ход цепи напоминает узор "меандр" (см. топологическую схему — плоскую развертку b-цилиндра — сбоку). На этой схеме b-тяжи изображены стрелками. "Меандр" получается потому, что соседние по цепи b-тяжи являются соседями на поверхности цилиндра; они связаны водородными связями (связь, существующая между крайними b-тяжами, изображена на плоской развертке маленькими черточками). Ретинол-связывающий центр находится у оси цилиндра. Сам ретинол окрашен в фиолетовый цвет. Цветная кодировка прослеживает ход цепи по b-цилиндру. Цифры на топологической схеме указывают порядок структурных сегментов в цепи.
Продольная упаковка (Рис.13-6б) характерна для листов, которые лишь скручены, как пропеллер, но не изогнуты дугой. Такую упаковку обычно называют b-сэндвичем. Торцы b-сэндвича прикрыты нерегулярными петлями (вы это видите на Рис.13-8). В некоторых b-сэндвичах крайние тяжи b-листов столь сближены (иногда они даже соединены несколькими водородными связями), что эта упаковка принимает форму цилиндра с малым наклоном b-участков к оси.
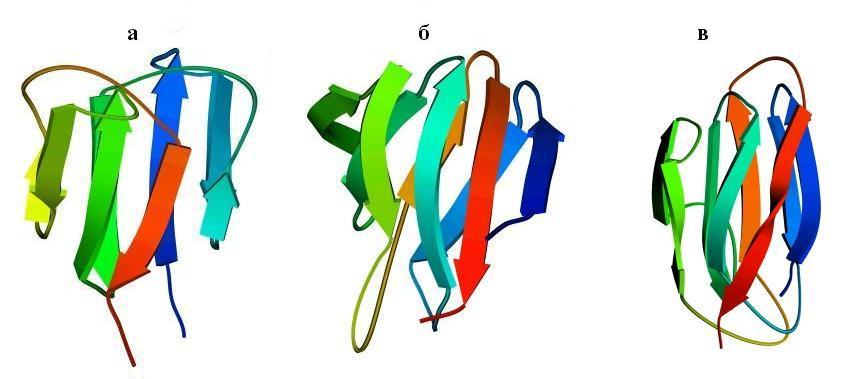
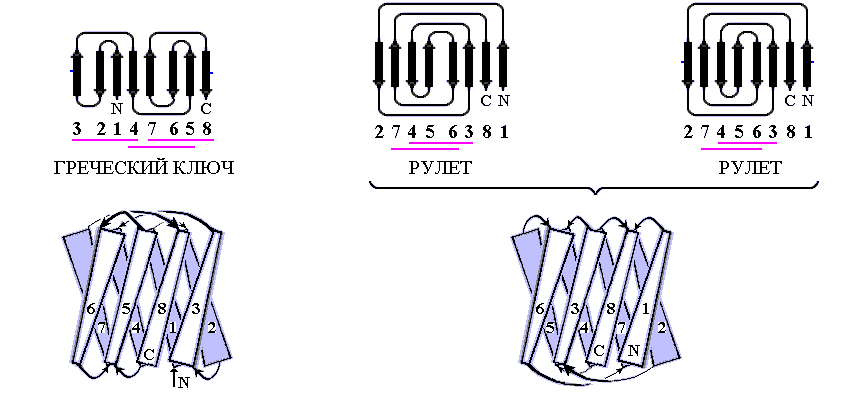
Рис.13-8. Примеры продольной упаковки b-листов. Мотивы укладки цепи в домене g кристаллина (а) (см. также Рис.13-2), в b-домене белка — катаболитического активатора (б) и в белке оболочки сателлитного вируса некроза табака (в). Внизу показаны топологические схемы этих белков. Все эти белки включают характерную топологию "греческого ключа" (или "согнутой пополам b-шпильки"), в которой четыре соседних по цепи b-тяжа антипараллельны, причем первый и четвертый связаны водородными связями. На топологических схемах двухслойный b-сэндвич развернут (как разрезанный сбоку цилиндр) и представлен в одной плоскости. Место разреза выбрано так, чтобы подчеркнуть симметрию укладки цепи. Так, в g кристаллине он приходится на стык b-тяжей 3 и 8 — с тем, чтобы подчеркнуть сходство структур первой (тяжи 1 — 4) и второй (тяжи 5 — 8) половинок домена. На топологической схеме сближением b-тяжей показаны Н-связи между ними; дополнительно, Н-связи между краями развертки (если они есть) показаны маленькими черточками на краях. Увеличенное расстояние между b-тяжами отделяет один b-лист сэндвича от другого (если там нет водородных связей b-тяжей). Домен g кристаллина содержит повтор, два греческих ключа: из тяжей 1 — 4 и 5 — 8; и еще один греческий ключ образован b-тяжами 4 — 7. Изображенные на рисунках (б, в) белки содержат "ключи" из тяжей 3 — 6 и 4 — 7. Более того, они имеют топологию многократно согнутой b-шпильки (она обычно называется "рулет"): здесь Н-связи соединяют также b-тяжи 1 и 8, 2 и 7, — в дополнение к связям тяжей 3 и 6, 4 и 5, типичным для "греческих ключей". Обратите внимание, что топология "греческого ключа" позволяет белковой цепи окружить ядро глобулы лучше, чем позволяет "меандр" (Рис.13-7): не только b-структурой с боков, но и петлями сверху и снизу. Обычно продольные упаковки b-листов представляют собой b-сэндвич (а, б), но некоторые из них (например — белка оболочки сателлитного вируса некроза табака и белков оболочки ряда других вирусов) можно представить также в форме b-цилиндров с колинеарными b-тяжами (в).
Необходимо подчеркнуть, что мотивов укладки белковой цепи имеется гораздо меньше, чем архитектур белковых глобул, а типов "штабелей" — гораздо меньше, чем мотивов укладки белковой цепи.
Так, одному и тому же "штабелю", т. е. одной и той же упаковке структурных сегментов могут соответствовать разные мотивы укладки цепи в глобулу, т. е. эти сегменты могут быть по-разному связаны единой полипептидной цепью. Рис.13-8 показывает, как один и тот же b-сэндвич (при различной топологии, т. е. при различном ходе цепи через этот сэндвич) служит основой структуры трех разных белковых доменов — домена g кристаллина (а), b-домена белка — катаболитического активатора (б), а также белка вирусной оболочки (в). Причем два последних белка имеют даже одинаковый мотив укладки цепи - то есть в них цепь одинаково проходит через одинаковый b-структурный штабель (это подчеркивается фигурной скобкой на рисунке 13-8)!
Здесь я не могу удержаться от искушения показать вам еще один белок, в основе которого лежит b-сэндвич. Это — иммуноглобулин, точнее — так (или примерно так) устроен каждый домен этого большого белка (Рис.13-9).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
