
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Более того, "шаблоны" можно применять и для описания кусков цепи, образующих более сложные структуры, — например, для описания b-a-b суперспиралей, состоящих из двух параллельных b-участков и a-спирали между ними (Рис.19-7). Такие структуры типичны для доменов, связывающих нуклеотиды. Особо важную роль в "шаблоне" играют так называемые "ключевые позиции", которые могут быть заняты только строго определенными аминокислотными остатками, — например, Gly: только этот остаток может находиться в конформации с f>60о, недоступной всем остальным остаткам.
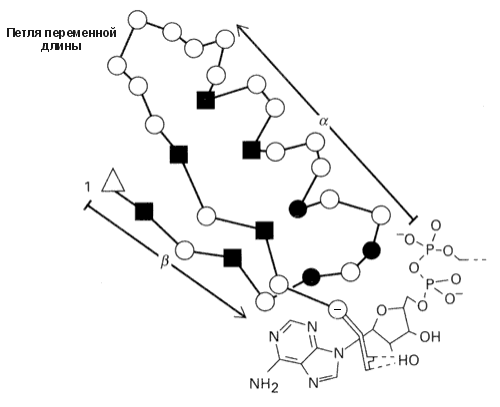
Рис.19-7. "Шаблон" суперспирали b-a-b, связывающей нуклеотиды. Квадратики отмечают ключевые позиции, обычно занимаемые сравнительно небольшими гидрофобными остатками (Ala, Ile, Leu, Val, Met, Cys): это — гидрофобное ядро суперспирали b-a-b. Черные кружки — позиции, занимаемые только Gly: здесь находятся резкие повороты цепи. Пустой треугольник отмечает первую позицию мотива b-a-b, где обычно находится положительно заряженная или дипольная боковая цепь. В последней (-) позиции мотива b-a-b находится аминокислота Asp или Glu, связывающая лиганд (нуклеотид). Рисунок, с небольшими изменениями, взят из R. K.Wierenga et al., J. Mol. Biol. (1986) 187:101-107.
Но вернемся к расчету вторичной структуры. Зная вклады отдельных взаимодействий в стабильность a-спирали, мы можем рассчитать свободную энергию спирализации любого участка цепи, а следовательно — и Больцмановскую вероятность образования спирали в любом месте полипептидной цепи, еще не свернувшейся в глобулу. Суммируя и усредняя эти вероятности, мы можем рассчитать и среднюю спиральность такого "несвернутого" полипептида. Вот уже более 15 лет мы используем для этого нашу программу ALB. В одном из режимов ("unfolded chain" — "развернутая цепь") она позволяет рассчитывать содержания a - и b-структуры в полипептидах и в несвернутых белковых цепях, — причем при разной температуре, ионной силе и рН раствора. Потом результат можно сравнить с опытными данными — например, с КД спектрами. Рисунок 19-8 показывает, что теоретический расчет неплохо совпадает с опытом.
| Рис.19-8. Теоретическая (вычисленная программой ALB — unfolded chain) и экспериментально найденная спиральность нескольких десятков пептидов при температуре 0о-5оС и разной ионной силе и рН раствора. Рисунок взят из A. V.Finkelstein, A. Y.Badretdinov & O. B.Ptitsyn, Proteins (1990) 10:287-299. |

Переходя к расчету и предсказанию вторичной структуры белков, глобулярных белков, необходимо учесть, что здесь к взаимодействиям, существующим в несвернутых цепях, добавляется взаимодействие каждого участка цепи с глобулой, строения которой мы не знаем. Точнее, мы не знаем ее детального строения, но знаем, что участки цепи как-то примыкают к гидрофобному ядру белка. В простейшем приближении взаимодействие с ядром можно аппроксимировать взаимодействием с "гидрофобным озером", на котором плавает белковая цепь (Рис.19-9).
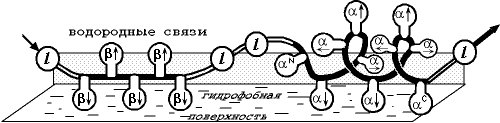
Рис.19-9. Флуктуирующая вторичная структура белковой цепи (здесь: b — b-тяж, l — петля, a — a-спираль) на поверхности гидрофобного озера, имитирующего белковую глобулу ("модель плавающих бревен"). Модель учитывает разное чередование обращенных к поверхности озера (![]() ), от поверхности (
), от поверхности (![]() ) и вдоль нее (®, ) боковых групп в разных вторичных структурах, а также эффекты на N - и С-концах спирали, объединенные и приписанные ее, соответственно, N - и С-концевым остаткам (aN, aC). Рисунок, с небольшими изменениями, взят из O. B.Ptitsyn & A. V.Finkelstein, Bipolymers (1983) 22:15-25.
) и вдоль нее (®, ) боковых групп в разных вторичных структурах, а также эффекты на N - и С-концах спирали, объединенные и приписанные ее, соответственно, N - и С-концевым остаткам (aN, aC). Рисунок, с небольшими изменениями, взят из O. B.Ptitsyn & A. V.Finkelstein, Bipolymers (1983) 22:15-25.
Зная из опыта силу гидрофобных взаимодействий, а из стереохимии a - и b-структуры — мотивы чередования в цепи боковых групп, глядящих в одну и ту же сторону и способных, следовательно, одновременно взаимодействовать с гидрофобной поверхностью, — мы можем сосчитать вероятность образования a-спирали и b-структуры в каждом месте белковой цепи. Этим также — но уже в режиме "глобулярная цепь" ("globular chain") — также занимается программа ALB (кстати, к ней можно обращаться по Интернет: http://indy. ipr. /~rykunov/alb/).
Вероятности рассчитываются при комнатной температуре. Почему при комнатной? Не лучше ли выделять только самое лучшее по энергии расположение a - и b-структур в цепи, т. е. рассчитывать все вероятности при 0оК?
Прежде всего, конечно, расчет относится к комнатной температуре потому, что все экспериментальные оценки стабильности, на которых базируется расчет, относятся к этой температуре.
Но еще важнее то, что вероятности, рассчитанные именно при такой температуре (300оК), лежащей чуть ниже нормальной температуры плавления белка (350оК) наилучшим образом позволяют отделить более вероятные a - и b-участки, в которых мы можем быть более или менее уверены, от тех "менее вероятных", в которых мы уверенными быть не можем.
Ведь, в сущности, мы стараемся предсказать вторичную структуру белка только по части тех взаимодействий, что ее держат в действительности: мы знаем (и то приблизительно) внутренние взаимодействия в этой структуре, но не знаем (или можем лишь крайне грубо оценить) те, что действуют между рассматриваемым куском цепи и остальной глобулой. А они очень мощны.
То есть мы находимся в том же положении, как если бы пытались предсказать, будет ли данный остаток внутри белка или на его поверхности, зная только его собственную гидрофобность — и больше ничего. И мы знаем, что такая задача имеет ответ, — но ответ не точный, а вероятностный. Этот ответ содержится в наблюдаемой статистике распределения остатков между нутром и поверхностью глобулы. Мы уже знаем, что она выглядит так:
ВЕРОЯТНОСТЬ_ВНУТРИ / ВЕРОЯТНОСТЬ_НА_ПОВЕРХНОСТИ ~ | (19.1) |
где TС — характерная температура белковой статистики, лежащая — как мы тоже помним — несколько ниже температуры плавления белка.
И — возвращаясь к предсказаниям белковых структур — по той же формуле (19.1), и с той же температурой ТС мы можем вероятностно предсказывать существование рассматриваемой вторичной структуры, зная только ее собственную свободную энергию.
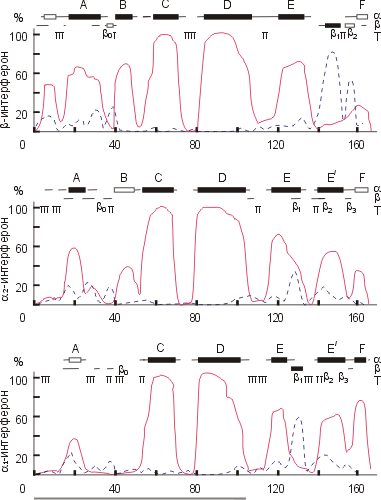
Рисунок 19-10 иллюстрирует расчет вторичной структуры в глобуле. Он был сделан в 1985 г. для интерферонов. Видно, что в них преобладают a-спирали, особенно в N-концевой части цепи, где (как было известно из других опытов) находится функциональный домен интерферона. В данном случае спирали эти предсказывались столь уверенно (а это бывает отнюдь не всегда) и столь одинаково в разных, не так уж и высоко гомологичных цепях, что мы попробовали — в том же 1985 г. — слепить домен из этих спиралей.
| Рис.19-10. Расчет вторичной структуры нескольких интерферонов. Абсцисса — номер остатка в белковой цепи, ордината — вычисленная вероятность его a-спирального ( ____ ) и b-структурного ( _ _ _ ) состояния. Сверху приведены предсказания наиболее вероятных a-спиралей (a), b-тяжей (b) и изгибов (Т). Черные прямоугольники — уверенное предсказание вторичной структуры, пустые — не столь уверенное предсказание, линии — данная вторичная структура в принципе не исключена, но маловероятна. Цепь, образующая N-концевой домен, подчеркнута внизу рисунка. Картинка взята из O. B.Ptitsyn, A. V.Finkelstein & A. G.Murzin, FEBS Letters (1985) 186:143-148. |
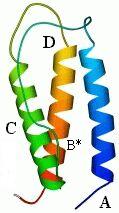
Полученный комплекс, пучок из трех больших спиралей и одной крошечной, показан на следующем рисунке (19-11а). Через пять лет после того, как было сделано это предсказание, была, наконец, получена рентгеновская структура интерферона b (Рис.19-11б), — и рентгеновская структура N-концевого его домена довольно точно совпала с той, что была предсказана нами.
|
|
|
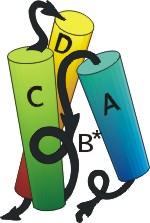
а б в
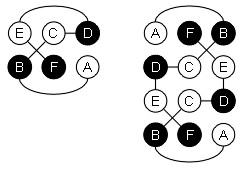
Рис.19-11. (а) Предсказание укладки цепи в N-концевом домене интерферонов, сделанное нами в 1985 г. [Ptitsyn, Finkelstein, Murzin, FEBS Letters (1985) v.186, p.143]. Три большие a-спирали (А, C, D) изображены цилиндрами; кроме того, была предсказана возможность существования отдельного спирального витка В. (б) Рентгенографическая структура N-концевого домена (С-концевой домен — не изображен) b-интерферона, расшифрованного в 1990 г. [Senda et al., Proc. Jpn. Acad. Sci., ser. B, Biopys. Biol. Sci. (1990) v.66, p.77]. Рисунок (б) дан в той ориентации, что и предсказанная модель (а), и на нем даны те имена больших спиралей (А, C, D), под которыми они значатся на рисунке (а). Район В* спиралеподобен, но не a-спирален в интерфероне b, однако в родственном ему интерфероне g там находится небольшая a-спираль [Ealick et al., Science (1991) v.252, p.698]. (в) Топологии b и g интерферонов по Ealick et al. Интерферон g состоит из двух субъединиц. Обратите внимание на "обмен" С-концевыми доменами [спиралями (E, F)] между этими двумя субъединицами.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
