
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
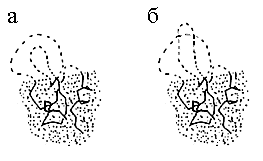
Рис.18-4. (а) Компактный интермедиат сворачивания белка с торчащими из него неупорядоченными петлями. Рост интермедиата соответствует смещению границы между фиксированной (глобулярной) и неупорядоченной (клубковой) частями белковой цепи. Успешное сворачивание интермедиата требует правильной заузленности петель в нем: полусвернутая структура с неправильным заузливанием (б) не может прорасти до правильно свернутого белка, ей сперва надо развернуться. Однако в цепи из ~100 звеньев может сформироваться лишь один-два узла, так что перебор интермедиатов с разной заузленностью петель не должен лимитировать скорость сворачивания белков.
Входящие в энтропию DS и энергию DE линейные и нелинейные члены должны войти и в свободную энергию DF=DE-TDS растущей глобулы. Однако, когда мы рассматриваем условия, в которых финальная глобула находится в термодинамическом равновесии с клубком (или почти в равновесии — скажем, когда она всего на пару RT стабильнее клубка), — то в разности DE-TDS большие (пропорциональные n) линейные члены взаимно аннигилируют. В самом деле, в самой точке равновесия DF=0 и в клубке (т. е. при n=0), и в финальной глобуле (т. е. при n=N). Значит, рост энтропии и падение энергии при последовательном сворачивании компенсируют друг друга в главном (линейном по длине свернутой цепи n) члене и, не будь поверхностных эффектов, DF было бы равно 0 на всем рассматриваемом пути сворачивания!
Следовательно, свободно-энергетический барьер связан только с относительно малыми нелинейными поверхностными эффектами. Значит, высота барьера на любом пути последовательного сворачивания (типа того, что изображен на Рис.18-2) пропорциональна не числу звеньев N (как то, фактически, полагал Левинталь), а всего лишь N 2/3 (Рис.18-3в).
В результате время достижения самой стабильной структуры растет с числом звеньев цепи N не "по Левинталю" (т. е. не как 10N и вообще не как экспонента от N), а всего лишь как exp(lN 2/3), — причем здесь наиболее важно то, что N 2/3 существенно меньше, чем N. В то же время тщательная оценка коэффициента l показывает, что l=1±0.5, причем конкретная величина l зависит как от формы укладки цепи в глобулу, так и от распределения сильно и слабо притягивающихся аминокислотных остатков в ней.
Причина того, что самая стабильная структура белка достигается всего за
ВРЕМЯ ~ exp(N 2/3) наносекунд, | (18.2) |
— в том, что падение энтропии по ходу последовательного сворачивания почти тут же компенсируется энергией возникающих взаимодействий.
Отметим, что на рассмотренном нами пути глобулярный зародыш белковой структуры не перестраивается по ходу сворачивания (на что потребовалось бы гигантское время), а все перестройки происходят только в рыхлом клубке (и потому — быстро).
Примечание. Не следует забывать, что мы рассмотрели сейчас только один из возможных сценариев сворачивания, и что не исключено, что рассмотрение других путей сворачивания белка может существенно понизить (но никак не повысить!) оценку времени его сворачивания, даваемую формулой (18.2). Кроме того, наша оценка относится к точке термодинамического равновесия клубка и нативной структуры белка, где экспериментально наблюдаемое время сворачивания этой структуры максимально, — оно может превосходить время сворачивания в нативных условиях на несколько порядков (см. ниже, Рис.18-6). Поэтому оценка (18.2) носит не столько количественный, сколько принципиальный характер, — она решает парадокс Левинталя и объясняет, почему белковая цепь может найти свою самую стабильную структуру не за астрономически огромное, а за биологически-разумное время.
Как бы то ни было, полученная оценка показывает, что цепь из 100 — 150 аминокислотных остатков должна находить свою самую стабильную структуру за секунды или минуты. Она объясняет также, почему большие белки состоят (согласно старому принципу "разделяй и властвуй") из отдельно сворачивающихся стабильных доменов — или, как теперь говорят в данном случае, "фолдонов" (от слова "fold" — "укладка"): иначе их сворачивание было бы слишком медленным.
Для очень длинных цепей, состоящих из многих тысяч звеньев, лимитирующим фактором мог бы стать "квази-Левинталевский" перебор по-разному заузленных интермедиатов — ведь узел в петле (ср. Рис.18-4а и 18-4б) нельзя распустить, не разрушив глобулярную часть интермедиата. Однако столь больших белков в природе нет (может быть, "потому и нет"), а в доменах — цепях из сотни-другой звеньев — много узлов быть не может (компьютерные эксперименты показали, для образования одного узла нужно порядка сотни звеньев цепи), так что перебор по-разному заузленных конформаций увеличит время достижения стабильной структуры домена всего в несколько раз.
До сих пор мы рассматривали сворачивание белковой цепи вблизи точки фазового перехода, когда только одна ("нативная") ее структура сравнима по стабильности с клубком, а все прочие, даже взятые вместе, нестабильны (Рис.18-5а).
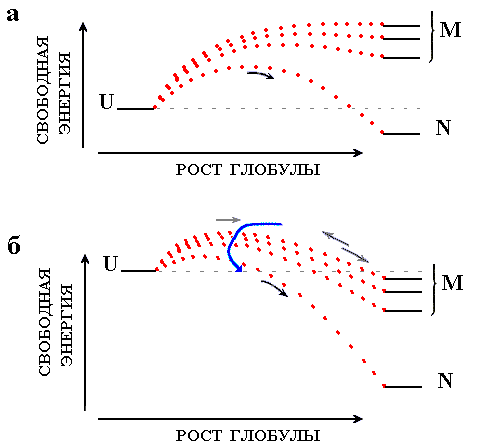
Рис.18-5. Сворачивание белка при разных условиях. Тонкие линии показывают уровни свободной энергии клубка (U), самой стабильной ("нативной") укладки цепи (N), и прочих ("misfolded", конкурирующих с самой стабильной) глобулярных структур (М). Точечные пунктирные линии показывают изменение свободной энергии по ходу сворачивания. Относительно небольшая их шероховатость (см. график DF на Рис.18-3) на данном рисунке не показана. Высшие точки этих пунктирных линий соответствуют свободным энергиям переходных состояний на соответствующих путях сворачивания. Основные сценарии процесса сворачивания: (а) Самая стабильная структура цепи N стабильнее клубка U, а все конкурирующие с ней структуры M в сумме менее стабильны, чем клубок. Быстрое и безошибочное сворачивание самой стабильной структуры. (б) Помимо структуры N, многие свернутые структуры M более стабильны, чем клубок U. Идет быстрое сворачивание многих "неправильных" структур с их последующим, очень медленным частичным разворачиванием и переходом в нативную структуру N. Стрелки показывают главное русло процесса самоорганизации стабильной пространственной структуры белка.
Что произойдет со скоростью самоорганизации цепи при удалении от точки фазового перехода? Нас, естественно, интересует случай, когда стабильность нативной структуры растет (когда ее стабильность падает — эта структура просто не образуется).
Рост стабильности нативной глобулы (например, при понижении температуры или при разбавлении раствора денатуранта водой) приводит сперва к увеличению скорости достижения нативной структуры, так как свободная энергия интермедиатов ее сворачивания падает, а конкурирующие с нативной структурой укладки цепи все еще остаются нестабильными (Рис.18-5а). Такое ускорение, действительно, наблюдается (см. подъем графика на Рис.18-6, слева) — до определенного предела (см. плечо графика на левом краю Рис.18-6). По-видимому, максимальная скорость сворачивания должна достигаться, когда конкурирующие с нативной структурой "неправильные" компактные укладки цепи (а их стабильность тоже растет при понижении температуры и разбавлении денатуранта) как раз сравниваются по стабильности с клубком.
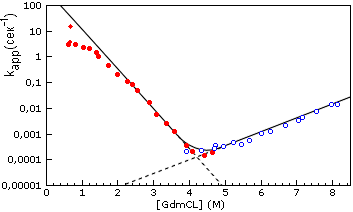
Рис.18-6. Зависимость скорости ренатурации и денатурации белка (лизоцима куриного яйца) от концентрации гуанидингидрохлорида. Экспериментальные точки изображают величину kapp = ku®N + kN®u, видимой характерной скорости приближения к равновесию между нативной и развернутой формами белка. Заполненные кружки получены при разбавлении раствора GdmCl, в котором изначально находился денатурированный белок, т. е. при полной или частичной ренатурации белка. При этом kapp » ku®N. Пустые кружки получены при добавлении GdmCl к изначально нативному белку, т. е. при полной или частичной его денатурации. При этом kapp » kN®u. Пунктир показывает экстраполяцию величин ku®N и kN®u в области излома шеврона. Загиб обеих линий экспериментальных точек в нижней части графика (при »4.5 М GdmCl) соответствует области термодинамического равновесия нативной и развернутой форм белка. В этих условиях время ренатурации составляет ~104 сек, — при том, что в "почти чистой" воде, при 0.6 М GdmCl (верхняя левая часть кривой), оно приближается к всего 0.1 сек. Картинка, с небольшими изменениями (добавлены экстраполяционные пунктиры) взята из T. Kiefhaber Proc. Natl. Acad. Sci. USA (1995) 92:9029-9033.
Область наиболее быстрого сворачивания исследовалась Шахновичем и его сотрудниками на простых компьютерных моделях белковых цепей. Эти работы как бы моделировали сворачивание белка в "нативных условиях". Они показали, что характерное время максимально быстрого сворачивания растет с длиной цепи много медленнее, чем то следует из формулы (18.2), оценивающей время сворачивания в точке термодинамического равновесия нативного и развернутого белка. А именно, компьютерный эксперимент показал, что время максимально быстрого сворачивания растет с числом звеньев цепи N вовсе не как exp(N 2/3), как то должно быть в точке равновесия, а как N 6 для "случайных" цепей и даже как N 4 для цепей, усиленно отобранных на предмет максимально быстрого сворачивания (и обладающих поэтому очень большой щелью между энергией самой стабильной структуры и энергиями прочих компактных укладок цепи).
Все это еще раз подчеркивает обсуждавшуюся на прошлой лекции сильную зависимость времени сворачивания белка от условий опыта, и прежде всего от стабильности нативной и конкурирующих с ней структур белка.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
