
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Обратите внимание на ряд существенных моментов.
Первое. Суммарный переход совершается на расстояние около 40![]() . Такое расстояние нельзя покрыть одним туннельным прыжком (такой прыжок занял бы ~ 1040x10- 15 сек ~ ~1025 сек ~ 1017 лет — на него не хватило бы всего времени жизни Вселенной). Однако конструкция белка разбивает этот большой прыжок на четыре малых — от одного притягивающего электрон пигмента к другому — и электрон покрывает эти 40
. Такое расстояние нельзя покрыть одним туннельным прыжком (такой прыжок занял бы ~ 1040x10- 15 сек ~ ~1025 сек ~ 1017 лет — на него не хватило бы всего времени жизни Вселенной). Однако конструкция белка разбивает этот большой прыжок на четыре малых — от одного притягивающего электрон пигмента к другому — и электрон покрывает эти 40![]() за долю миллисекунды.
за долю миллисекунды.
Второе. Для того, чтобы электрон не вернулся бы тотчас со второго пигмента на первый, а пошел бы дальше, на третий и т. д. — его суммарная (потенциальная + кинетическая) энергия должна падать по ходу процесса — или, иными словами, электрон должен на каждом шаге переходить с высокоэнергетической атомной орбиты на низкоэнергетическую. Конструкция фотосинтетического реакционного центра призвана обеспечить такое понижение энергии электрона на пигментах по ходу процесса.
Третье. Совершая туннельный переход, электрон не расходует энергии на преодоление барьера (здесь нет никакого "трения") — однако электрон на каждом шаге идет от более высокоэнергетической атомной орбиты к более низкоэнергетической, его энергия понижается — и она расходуется на то, чтобы сделать туннельный переход "эффективным", т. е. необратимым.
И последнее. Туннельный (или, как говорят, "под-барьерный") переход можно отличить от обычного активационного механизма перехода через энергетический барьер по тому, что на скорость туннельного перехода температура не влияет, а скорость активационного перехода (пропорциональная exp(-DE#/kBT), где DE# — энергия активационного барьера, а Т — температура) резко уменьшается при падении температуры.
Лекция 13
Теперь в фокусе нашего внимания будут находиться глобулярные, а точнее — водорастворимые глобулярные белки. Именно они наиболее исследованы: для сотен — изучена спонтанная самоорганизация, для тысяч — расшифрована атомная трехмерная структура. Поэтому именно их обычно имеют в виду, говоря о "типичных белковых структурах", о "закономерностях, наблюдаемых в строении и самоорганизации белков" и т. д. После этой необходимой оговорки — обратимся к структурам глобулярных белков.
Рентгенструктурные исследования (а позже, — двух - и многомерный ЯМР) позволили установить — за 40 лет интенсивной работы многих десятков лабораторий — атомную структуру порядка 2000 белков (а если считать и мутантные формы, и различные функциональные состояния — то раз в пять больше).
Что можно увидеть в структурах белков — водорастворимых глобулярных белков — с птичьего полета?
Мы видим, что небольшие (из 50 — 150, реже из 200 — 250 остатков) цепи укладываются в компактную глобулу диаметром 25-40![]() (Рис.13-1); и что более крупные белки состоят из нескольких таких субглобул — "доменов" (Рис.13-2). Белковая цепь упаковывается в глобулу так же плотно, как органические молекулы — в кристалл. Это видно и при взгляде на поверхность белка (Рис.13-1а), и на "срезе" белковой глобулы, показанном на Рис.13-1б. Однако, рассматривая белок, нам предстоит рассматривать не плотноупакованные электронные облака (или Вандерваальсовы поверхности) атомов — иначе ничего не будет видно внутри белка — а только очищенные от "мяса" атомов (Рис.13-1в) и даже от боковых групп вообще (Рис.13-1г) скелеты белковых молекул. Поэтому не поддавайтесь часто создаваемому рисунками ощущению "рыхлости" белковых глобул!
(Рис.13-1); и что более крупные белки состоят из нескольких таких субглобул — "доменов" (Рис.13-2). Белковая цепь упаковывается в глобулу так же плотно, как органические молекулы — в кристалл. Это видно и при взгляде на поверхность белка (Рис.13-1а), и на "срезе" белковой глобулы, показанном на Рис.13-1б. Однако, рассматривая белок, нам предстоит рассматривать не плотноупакованные электронные облака (или Вандерваальсовы поверхности) атомов — иначе ничего не будет видно внутри белка — а только очищенные от "мяса" атомов (Рис.13-1в) и даже от боковых групп вообще (Рис.13-1г) скелеты белковых молекул. Поэтому не поддавайтесь часто создаваемому рисунками ощущению "рыхлости" белковых глобул!

Рис.13-1. Картинки, изображающие строение небольшого белка — a-субъединицы интерлейкина 8 — при разной степени схематизации. (а) Атомная модель (изображены только "тяжелые", не-водородные атомы: синие — азоты, красные — кислороды, серые — углероды); в силу того, что цепь в глобуле упакована плотно, мы видим лишь поверхность белка. (б) Срез атомной модели подчеркивает плотность упаковки. (в) Скелетная модель главной цепи (темная линия) и боковых групп (более светлые отростки). (г) Ход главной цепи. (д) Схема строения белка, на которой выделены вторичные структуры (две a-спирали и один b-лист из трех b-тяжей) в главной цепи белка. (е) Структурный каркас белка, сложенный из вторичных структур. Все рисунки даны в одной и той же проекции и в одинаковом масштабе.
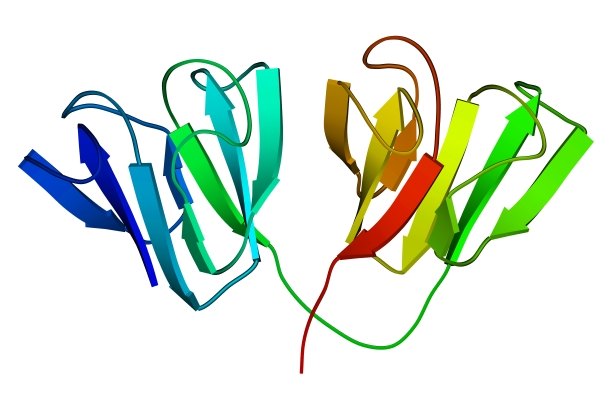
Рис.13-2. Глобулярные домены в g-кристаллине. Цветная кодировка трассирует ход цепи (от синего на N-конце цепи к зеленому в середине и к желтому и красному на С-конце).
Каркас пространственной структуры подавляющего большинства глобул (доменов) сложен из уже изученных нами регулярных вторичных структур — a-спиралей и b-листов (Рис.13-1д), которые стабилизованы регулярными водородными связями в регулярной главной цепи. Кстати, эти вторичные структуры были теоретически предсказаны Полингом, Кори и Брэнсоном еще до расшифровки атомного строения белковых молекул. "Штабель", сложенный из этих структур (Рис.13-1е), определяет основные особенности строения белка.
Каркас из a - и b-структур как бы окружает гидрофобное ядро (или ядра) белка, а нерегулярные петли лежат дальше, на периферии глобулы. Петли практически никогда не входят внутрь белковой глобулы — и неудивительно: их не вовлеченные во вторичную структуру пептидные группы не должны порывать своих водородных связей с водой, это вело бы к нестабильности глобулы.
В зависимости от строения каркаса, глобулярные белки подразделяются на: "чистые" b белки; "чистые" a белки; и "смешанные" a/b, a&b и a+b белки. Строго говоря, эта классификация типов белковых структур относится к малым белкам, а также к отдельным доменам (т. е. к компактным субглобулам, из которых сложены большие белки), так как большие белки могут содержать, например, и b-, и a-домены одновременно.
Нас будет прежде всего интересовать: 1) архитектура упаковки a - и b - структурных сегментов в компактную глобулу (Рис.13-1е); и 2) ход цепи через глобулу (Рис.13-1д), — или, как часто говорят, "топология белковой глобулы".
При этом мы часто будем использовать упрощенные схемы строения белков (Рис.13-3). Такое упрощение достигается не только обращением основного внимания на вторичные структуры (при пренебрежении к детальному строению петель), но и пренебрежением различий в размерах этих структур и к деталям их взаимной ориентации [при этом мы переходим от "укладок" ("folds", Рис.13-3а) к "мотивам укладок" ("folding patterns", Рис.13-3б) белковых цепей].
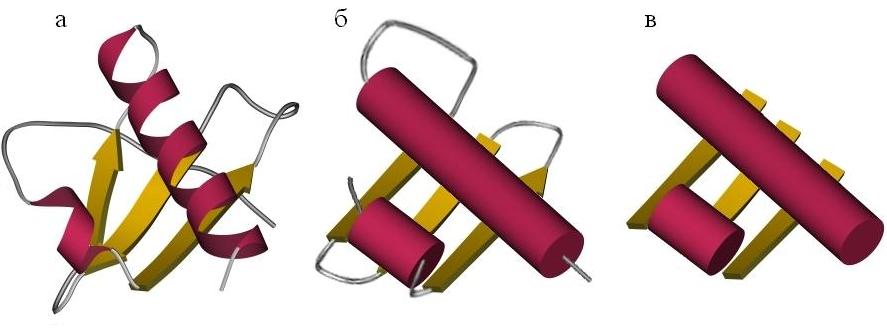
Рис.13-3. Упрощенные представления белковых структур. (а) Детальная укладка ("fold"), описывающая размещение вторичных структур в цепи белка и в пространстве (см. также Рис.13-1д). (б) Мотив укладки белковой цепи ("folding pattern"): опущены детали хода петель, размера и точной ориентации a-спиралей (изображенных в виде параллельных цилиндров) и b-тяжей (изображенных в виде стрелок). (в) Упаковка: штабель ("stack") структурных сегментов: петли удалены, опущены размер, ориентация и направление a-спиралей и b-тяжей (изображенных поэтому в виде лент).
Упрощение это оправдывается тем, что детали строения петель и точные размеры и ориентации структурных сегментов (и даже некоторые маленькие структурные сегменты) не сохраняются, когда мы переходим от рассматриваемого белка к его довольно близкому (имеющему сходную аминокислотную последовательность, т. е. явно общее происхождение) родственнику — например, от гемоглобина a к гемоглобину b (Рис.13-4).
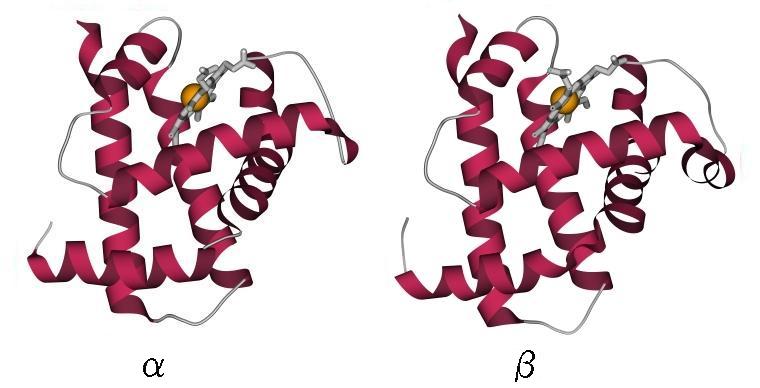
Рис.13-4. Два близко родственных белка: гемоглобин a и гемоглобин b лошади (оба — с гемом). Найдите сходства и различия! (Подсказка: при общем высоком сходстве — различия в деталях конформации петель, в деталях ориентации некоторых спиралей, и в одном дополнительном спиральном витке в b глобине, справа).
На следующем, более высоком уровне упрощения, необходимом для классификации структур белковых глобул, мы иногда вообще будем рассматривать лишь упаковку структурных сегментов в глобулу — т. е. сложенные из вторичных структур упаковки, или "штабели" ("stacks"), — временно забыв о петлях, соединяющих эти вторичные структуры в единую молекулу (Рис.13-3в).
Я буду специально пользоваться такими упрощенными схемами мотивов укладки и упаковки белковой цепи, — вырисовывая в то же время, для сравнения, и "истинные" структуры белков так, как их дает компьютер. Казалось бы, зачем нужны упрощенные схемы, если компьютер может нарисовать "все как есть". Дело в том, однако, что "все как есть" содержит очень много деталей, а схемы берут из них главные, повторяющиеся в разных сходных белках. Поэтому они удобны и для классификации белковых структур, и для выделения их главных, типических черт. Рассматривая подробные картинки белков, вы все равно в уме выделяете такие типические черты, — а схемы просто позволяют вам систематизировать ваше интуитивное выделение. Кроме того, они дают возможность составить "словесный портрет" белка. На практике такие схемы и такие словесные портреты вам понадобятся сразу же, как только вы захотите выяснить, на кого похож интересующий вас белок: они подчеркивают главное, убирая детали. Конечно, эти "детали" могут быть самыми важными для функционирования белка, и мы с этим встретимся, — но это обстоятельство только подчеркивает относительную независимость функции белка от мотива укладки цепи в нем.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
