
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
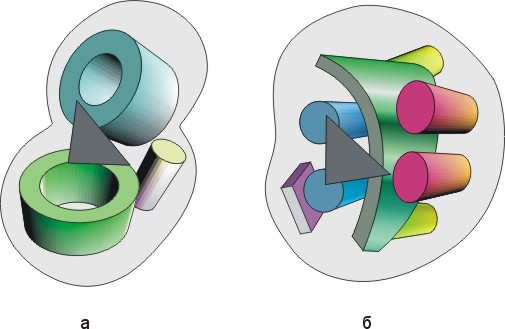
Рис.20-16. Схема строения сериновых протеаз типа трипсина (а) и субтилизина (б). На фоне общего контура глобул показаны a-спирали, b-листы и b-цилиндры. Район активного центра показан черным треугольником.

Рис.20-17. Схема строения каталитических центров сериновых протеаз типа трипсина (а) и субтилизина (б). Черным выделены фрагменты главной цепи (вплоть до Сb-атомов); направление хода цепи в этих фрагментах показано стрелками. Кружком показано место оксианионовой дыры. Видно, что только расположение концов боковых групп каталитической триады переноса заряда (Ser, His, Asp) и — с натяжкой — положение оксианионовой дыры инвариантно в этих двух классах сериновых протеаз, в то время как кардинальные отличия наблюдаются даже в ходе главной цепи у ключевых каталитических остатков His и Asp.
Более того: существуют протеазы, совершенно не похожие на сериновые протеазы даже в зоне каталитического центра. Я имею в виду, в частности, металлопротеазы типа карбоксипептидазы или термолизина. У них ключевую роль в катализе играет плотно встроенный в белок ион Zn++. Имея маленькие радиусы, многозарядные металлические ионы создают вокруг себя очень сильное электрическое поле. Это позволяет им — в металлопротеазах — активировать молекулу воды (отщепляя от нее Н+) с тем, чтобы та осуществила бы далее разрыв пептидной связи (ср. Рис.20-10), и одновременно — стабилизировать зарядом Zn++ отрицательный заряд на тетраэдрическом переходном состоянии разрываемого пептида.
Итак, одна и та же химическая реакция может осуществляться совсем разными белками. При этом одну и ту же ключевую роль — роль мощных электрических резаков — в одних белках играют многозарядные металлические ионы, в других — легко принимающие электроны или протоны органические кофакторы, в третьих — активированные своим окружением (как в трипсине) боковые группы...
С другой стороны, как мы помним, разные белки с одной и той же характернейшей (Рис.20-7) формой a/b-цилиндра (а равно и белки, имеющие форму укладки Россманна) могут катализировать совершенно разные реакции.
Все это показывает довольно широкую независимость общего строения белка от его ферментативной активности.
До сих пор я почти ничего не говорил о конформационных изменениях белковых молекул в ходе их функционирования — и не случайно: пока речь идет — как у нас до сих пор — о катализе одной, отдельно взятой энзиматической реакции — конформационная гибкость белка может только ухудшить его каталитические свойства. В самом деле, для эффективного катализа нужно преимущественное связывание именно переходного состояния (Рис.20-11), — в пику отличающимся от него лишь на какой-то ангстрем начальному и конечному состояниям, — и это преимущественное связывание нацелено на синтез или разрыв очень жестких ковалентных связей. Так что рвать их "гибким" белком — все равно, что резать проволоку подушкой...
Впрочем, здесь надо добавить следующее. Когда активный центр белка не идеально подогнан под свою функцию — а идеальность подгонки требует сверхтщательной работы естественного отбора — этот центр может требовать небольшой деформации для перехода в активную форму. И тогда такая деформация будет, действительно, функционально необходима — она будет, время от времени, доводить несовершенный белок до кондиции. Однако здесь надо четко отличать нужду (деформацию, искупающую несовершенство геометрии активного центра) от добродетели (от совершенства белка).
Конечно, даже самый лучший белок несколько деформируется в процессе работы — ведь он не алмазный — но это играет примерно ту же функциональную роль, что деформация бумаги под пером: чем меньше — тем лучше.
Другое дело, когда белок должен перейти от одного действия к другому: тогда его деформация, обеспечивающая переход от одной роли к другой, действительно, играет важную функциональную роль.
Так, в многих белках проникновение субстрата в активный центр требует небольшого расхождения близких к этому центру боковых групп. Однако, после того, как субстрат сел на этот центр, его каталитическая работа движения этих групп уже не требует.
Можно сказать, что подвижность нужна при подготовке к реакции, а жесткость — в момент ее проведения. Это похоже на военное "правило Клаузевица" — "на марше — отдельно, в бою — вместе".
Мы уже познакомились с одной функционально-важной деформацией на примере регуляции ДНК-связывающей активности trp-репрессора. Сейчас мы рассмотрим вопрос о функционально-важных деформациях более внимательно.
Прежде всего посмотрим, как деформация белка помогает сочетать стадии цикла СВЯЗАТЬ ® ТРАНСФОРМИРОВАТЬ ® ОТПУСТИТЬ. Рисунок 20-18 демонстрирует индуцированное соответствие (введенный Д. Кошландом термин) фосфорилирующего белка — гексокиназы — к его субстратам. Этот белок переносит фосфатную группу с АТФ на глюкозу. Но — в принципе, по химии реакции — эта же фосфатная группа может быть перенесена и на воду; однако этого не происходит. В попытке ответить на вопрос, почему этого не происходит, Кошланд постулировал следующее. 1) До связывания с субстратом фермент находится в "открытой" форме (в которой он может захватить субстрат из воды, но не способен провести его фосфорилирование). 2) После связывания с субстратом он — фермент — переходит в "закрытую", каталитически-активную форму, где все части активного центра собраны воедино и способны катализировать реакцию фосфорилирования, но вода вытеснена из активного центра и потому не конкурирует с субстратом за фосфорилирование. 3) После каталитического акта фермент снова открывается, и фосфорилированный субстрат уходит.
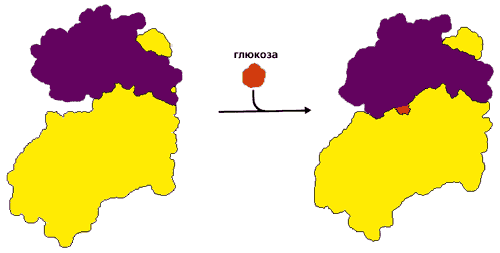
Рис.20-18. Индуцированное соответствие при функционировании гексокиназы. В открытой форме два домена разделены глубокой щелью, куда может заплыть глюкоза. Когда глюкоза попала в щель, домены поворачиваются, щель закрывается, вода из нее вытесняется, а все компоненты каталитического центра сходятся вместе. Картинка взята из [5].
Впоследствии опыт полностью подтвердил эту гипотезу (Рис.20-18), — но только для тех белков, которым нужно скрыть обрабатываемый субстрат от конкурирующей с ним воды. Для действия трипсина, например, этого не нужно, — и в нем индуцированного соответствия субстрату не наблюдается: трипсин (а также — химотрипсин, эластаза, субтилизин и т. д.) не деформируется и опознает субстрат по простейшему принципу "ключ — замок".
Хочу привлечь ваше внимание к тому, что индуцированное соответствие достигается смещением либо крупных блоков (Рис.20-3), либо целых белковых доменов (Рис.20-18), — а не полной перестройкой укладки белковой цепи. А смещения эти происходят в основном путем мелких локальных деформаций. [Аналогия: мышцы сокращаются ("локальная деформация"), и пальцы ("домены") сжимаются в кулак.]
То же самое имеет место и во всех других известных случаях "конформационных перестроек белков" — за немногими исключениями, когда из глобулы иногда вырывается и переходит в нерегулярную конформацию целая a-спираль или b-тяж. Одна из самых больших известных мне перестроек происходит в калмодулине. Сам по себе он имеет форму гантели, "головки" которой — a-спиральные домены — разнесены на большое расстояние "ручкой" — одной длинной a-спиралью. Однако при связывании калмодулина с рядом других белков "головки" бывшей гантели, не ломаясь, сближаются и слипаются друг с другом и с белком-целью, а бывшая "ручка" — длинная a-спираль — разрушается.
Относительная автономия доменов хорошо видна в большом семействе белков, называемых дегидрогеназами. Эти белки катализируют сходные реакции типа окисления алкогольных групп при помощи кофактора НАД, легко принимающего оторванные протоны. Однако окисляют они разные вещества: алкогольдегидрогеназа — спирт (этанол), лактатдегидрогеназа — лактат, и т. д.
Дегидрогеназы состоят из двух доменов (Рис.20-19), соединенных перемычкой.
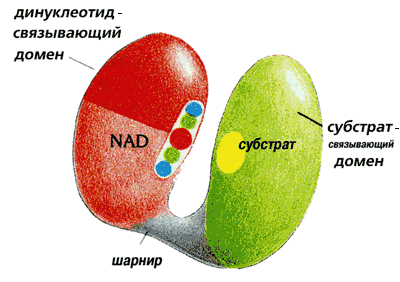
Рис.20-19. Цепи НАД-зависимых дегидрогеназ свернуты в два отдельных домена, соединенных относительно гибкой перемычкой. Один из них более или менее универсален (т. е. очень сходный домен встречается в разных белках) — он связывает кофактор НАД. Второй — не универсальный, а индивидуальный в каждой дегидрогеназе домен, — связывает обрабатываемый субстрат. Картинка взята из [5].
Один, связывающий обрабатываемый субстрат домен, индивидуален в каждой из дегидрогеназ. Например, в алкогольдегидрогеназе он содержит b-цилиндр, а в глицеральдегид-3-фосфат дегидрогеназе — плоский b-лист.
В то же время второй из этих доменов — тот, что связывает кофактор НАД — у всех НАД-зависимых дегидрогеназ устроен практически одинаково, хотя в одних дегидрогеназах он находится в N-, а в других — в С-концевой половине цепи, и несмотря на отсутствие видимой гомологии в первичных структурах НАД-дегидрогеназ. В этом a/b домене цепь обладает "укладкой Россманна" (см. Рис.20-7б), причем сходство распространяется и на мелкие детали НАД-связывающих доменов разных дегидрогеназ: большую часть этих доменов можно наложить друг на друга с точностью до 2 ![]() , — включая и место связывания НАД [это, по-видимому, показывает, что пространственная структура лучше помнит родство белковых доменов, чем первичная.]
, — включая и место связывания НАД [это, по-видимому, показывает, что пространственная структура лучше помнит родство белковых доменов, чем первичная.]
Таким образом, разнообразие действия дегидрогеназ обеспечивается — при универсальности НАД-связывающего домена, несущего каталитический центр, — разнообразием субстрат-связывающих центров, расположенных на "субстрат-связывающих" доменах, устроенных по-разному: в структуре целого белка эти две части активного центра соприкасаются (Рис.20-19).
В заключение я хочу рассказать о неконтактном, — или, как говорят, аллостерическом взаимодействии активных центров. Аллостерические взаимодействия между различными связывающими и активными центрами, особенно в олигомерных белках, играют важнейшую роль в контролировании и интегрировании биохимических реакций. Сейчас я рассмотрю (в самой упрощенной форме) только один, наиболее изученный аллостерический белок — гемоглобин.
Это — тетрамер, точнее — комплекс из двух "a" и двух похожих на них "b" цепей (Рис.20-20а). Его дело — связать кислород в легких (где его много), донести до мышц (где его мало) и отдать мышечному миоглобину, мономерному белку, похожему на любую из четырех субъединиц гемоглобина. Активный центр миоглобина и каждой a и b субъединицы гемоглобина — гем: кофактор, связывающий одну молекулу О2 (Рис.20-20б). И успешность действия гемоглобина как транспортера О2 определяется именно аллостерическим, в данном случае — концертным взаимодействием четырех гемов в нем.
а б
б 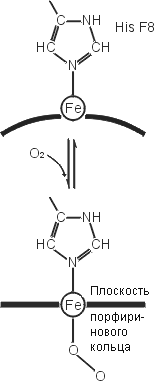
Рис.20-20. (а) Общая структура гемоглобина, состоящего из двух a и двух b цепей; каждая из четырех цепей окрашена в свой цвет, гемы показаны фиолетовыми скелетными моделями с оранжевым ионом железа внутри. (б) Схема связывания кислорода гемом в гемоглобине. До связывания О2 ион железа (Fe++) гема находится в высоко-спиновой форме, в которой два внешних электрона находятся на двух разных орбитах. При этом диаметр Fe++ чуть великоват для того, чтобы войти в центр порфиринового кольца гема (которое при этом находится в слегка изогнутой форме). Связав О2, ион Fe++ приобретает низкоспиновую, чуть более компактную форму, в которой электроны с противоположными спинами занимают одну орбиту. Теперь Fe++ может войти в центр порфиринового кольца (которое при этом обретает плоскую форму). Показан также гистидин a-спирали F, координирующий ион железа. Картинка (б) взята из [9].
Связав кислород, ион железа гема входит в центр порфиринового кольца гема. При этом он чуть смещается и тянет за собой координирующий его гистидин a-спирали F. Это смещение — посредством множества мелких деформаций белка — слегка изменяет обводы связавшей О2 субъединицы. В тетрамерном гемоглобине эта субъединица — в такой форме — начинает деформировать (придавая им ту же внешнюю форму и тем наводя в них те же внутренние смещения атомов) другие, еще не связавшие О2 субъединицы гемоглобина. Теперь ион железа в них может уже легче связать кислород. В результате, связь одной молекулы О2 с гемоглобином провоцирует связывание еще трех О2. Точно так же, утеря одного О2 ведет к утере их всех.
То есть гемоглобин ведет себя как белок, связывающий четыре молекулы О2 одновременно, — и это отражается в нелинейной, S-образной зависимости насыщения этого (тетрамерного!) белка кислородом от его (О2) концентрации в крови (Рис.20-21). А у (мономерного!) миоглобина зависимость насыщения кислородом от концентрации О2 в крови S-образного прогиба не имеет.
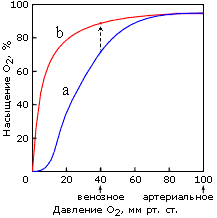
Рис.20-21. Кривые насыщения кислородом тетрамерного белка гемоглобина (а) и мономерного миоглобина (b). Отмечено артериальное давление кислорода, при котором он поглощается гемоглобином крови в легких, и венозное давление, при котором он отдается гемоглобином мышечному миоглобину. Экспериментальная кривая связывания О2 с миоглобином соответствует реакции первого порядка, а экспериментальная S-образная кривая связывания О2 с гемоглобином — реакции примерно третьего (а не четвертого, как для простоты сказано в тексте) порядка. Стрелка соответствует переходу кислорода с гемоглобина на миоглобин. Картинка, с небольшими изменениями, взята из [1].
Поэтому в легких, где кислорода много, гемоглобин им насыщается. А в тканях, где (венозное) давление О2 относительно мало, тетрамерный гемоглобин уже отдает О2, а мономерный миоглобин еще его хватает, — и передает О2 дальше по окислительной цепи мышц, — например, тех, что расширяют и сжимают легкие.
И так мы дышим и живем.
Контрольные Вопросы
ВВЕДЕНИЕ
1. Основные функции белков. Аминокислотная последовательность определяет пространственную структуру, пространственная структура — функцию. Обратное — неверно. Глобулярные, фибриллярные и мембранные белки.
2. Первичная, вторичная, третичная, четвертичная структура белка. Биосинтез белка; сворачивание белка in vivo и in vitro. Пост-трансляционные модификации.
ЭЛЕМЕНТАРНЫЕ ВЗАИМОДЕЙСТВИЯ
3. Стереохимия L-аминокислотных остатков. Валентные связи и углы между ними. Пептидная группа. Транс— и цис-пролины.
4. Вандерваальсово взаимодействие. Карты Рамачандрана для разрешенных конформаций аминокислотного остатка (глицин, аланин, валин, пролин).
5. Влияние водного окружения. Электростатические взаимодействия. Водородные связи. Гидрофобные взаимодействия.
6. Свойства боковых групп аминокислотных остатков. Неполярные, короткие полярные и длинные полярные боковые группы; заряженные группы; цистеины.
ВТОРИЧНЫЕ СТРУКТУРЫ ПОЛИПЕПТИДНЫХ ЦЕПЕЙ
7. Вторичная структура полипептидов. Спирали: 27, 310, a, poly(Pro). Антипараллельная и параллельная b-структура. b-изгибы.
8. Стабильность a-спирали в воде. Размер кооперативного участка. Переход спираль-клубок. Стабильность b-структуры. Скорость образования b-структуры.
ПРОСТРАНСТВЕННОЕ СТРОЕНИЕ БЕЛКОВ
9. Фибриллярные белки, их функции и их вторичные структуры: a-кератин, b-фиброин шелка, коллаген.
10. Мембранные белки, особенности их строения и функции. Бактериородопсин, фотосинтетический центр, порин. Каналы. Туннельный эффект.
11. Глобулярные белки. Упрощенное представление белковых структур. a - и b-слои. Редкость перекрывания петель и параллельности соседних по цепи структурных сегментов.
12. Строение b-белков: продольная и перпендикулярная упаковка b-листов, преимущественная антипараллельность b-структуры в b-белках. Правопропеллерность b-структурных листов.
13. Строение a-белков. Пучки и слои спиралей. Модель квазисферической глобулы из a-спиралей.
14. Строение a/b белков: параллельный b-слой, прикрытый a-спиралями. Строение a+b белков. Домены в белках.
КОДИРОВАНИЕ БЕЛКОВОЙ СТРУКТУРЫ АМИНОКИСЛОТНОЙ ПОСЛЕДОВАТЕЛЬНОСТЬЮ
15. Физические принципы строения белковой глобулы. "Стандартные" третичные структуры. Влияние формы структуры на число мутаций, не нарушающих структуру белка, или: Почему одних структур много, а других — мало?
16. Самоорганизация белков. "Парадокс Левинталя" и его решение. Нуклеационный механизм сворачивания. Интермедиаты и ловушки. Роль шаперонов.
17. Представление о подходах к предсказанию вторичных и пространственных структур белков по их аминокислотным последовательностям, и к "опознаванию" этих структур по гомологии последовательностей. "Шаблоны".
18. Белковая инженерия и дизайн.
КООПЕРАТИВНЫЕ ПЕРЕХОДЫ В БЕЛКОВЫХ МОЛЕКУЛАХ
19. Элементы термодинамики, статистической физики и химической кинетики. Что такое энтропия? Температура? Свободная энергия? Химический потенциал? Фазовый переход? Критерий Вант-Гоффа для перехода "все-или-ничего".
20. Понятие о теории абсолютных скоростей реакций. Нуклеация фазовых переходов 1-го рода, ее скорость. Кооперативные не-фазовые переходы.
21. Термодинамика де - и ренатурации. Как выглядит денатурированный белок? Клубок и расплавленная глобула.
22. Почему денатурация глобулярного белка — переход типа "все-или-ничего"? Распад плотной упаковки ядра белка и раскрепощение боковых групп. Проникновение растворителя в денатурированный белок.
ФУНКЦИОНИРОВАНИЕ БЕЛКОВ
23. Биохимическая функция белка и его структура. Активный центр — "дефект" глобулярной структуры. Каталитический и субстрат-связывающий центры; гидрофобный "карман". Кофакторы. Многовалентные ионы.
24. Механизм ферментативного катализа. Пример: сериновые протеазы. Теория переходного состояния в катализе и ее подтверждение методами белковой инженерии.
25. Узнавание "ключ-замок" и индуцированное соответствие. Ингибирование. Аллостерия — взаимодействие активных центров. Гемоглобин и миоглобин. Доменная структура: киназы, дегидрогеназы, иммуноглобины.
* * * * *
Заключение
Предложенный вашему вниманию курс основан на лекциях, которые читались мной в Пущинском университете и затем в Пущинском филиале МГУ в 1992-99 гг., а еще раньше, на московском Физтехе, — сначала моим учителем , а затем мной. Но на Физтехе лекции читались для физиков, а нынешние — для биологов. Поэтому их пришлось не только сильно обновить (наука не стоит на месте), но и кардинально переработать — для биологической аудитории.
Как я уже говорил, эти лекции отражают мои личные вкусы и пристрастия, мой личный круг научных интересов. Поэтому я буду благодарен за все вопросы и замечания, которые помогут мне лучше удовлетворить научные интересы аудитории.
24/IV/1999
Новая редакция: 24/V/2000

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
