
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
PРАСПЛ = exp[-(E'-TS')/kT] / { exp[-(E-TS)/kT] + exp[-(E'-TS')/kT] } = 1 / { exp[(DE-TDS)/kT] + 1 }, | (16.1) |
где DE=E'-E, DS=S'-S. Вероятность пребывания "единицы плавления" в твердом состоянии есть (для перехода "все-или-ничего") PТВ = 1-PРАСПЛ. Производная dPРАСПЛ/dT показывает, как быстро меняется PРАСПЛ с температурой. Простое вычисление (с использованием уже известной нам формулы d(F/T)/dT = d[(E-TS)/T]/dT = - E/T2) показывает, что
dPРАСПЛ/dT = PРАСПЛ (1-PРАСПЛ) (DE/kT2). | (16.2) |
Пожалуй, надо расписать здесь все преобразования подробнее, чтобы не принимать на веру, а понимать. Обозначим (DE-TDS)/kT через X. Тогда dX/dT = - DE/kT2, PРАСПЛ = 1/(eX+1), PТВ = 1-PРАСПЛ = eX/(eX+1), и
dPРАСПЛ/dT = d[1/(eX+1)]/dT = - [1/(eX+1)2] x [deX/dT] = - [1/(eX+1)2] x eX x [dX/dT] =
= - [1/(eX+1)] x [eX/(eX+1)] x [dX/dT] = PРАСПЛ (1-PРАСПЛ) x (-dX/dT) = PРАСПЛ (1-PРАСПЛ) (DE/kT2)
Точка середины перехода — точка самого быстрого изменения PРАСПЛ, т. е. максимум производной dPРАСПЛ/dT — падает на температуру Т0, где PРАСПЛ=PТВ = 1/2. В этой точке величина PРАСПЛ x (1-PРАСПЛ) проходит через максимум, равный 1/4. Здесь наклон кривой PРАСПЛ(Т) максимален и равен
(dPРАСПЛ/dT)|T=To = (1/4) DE/kT02. | (16.3) |
Графическое определение ширины перехода показано на Рис.16-3, слева. Оно делается путем линейной экстраполяции максимального наклона кривой PРАСПЛ(Т) до пересечения с базовыми прямыми, соответствующими нативному (PРАСПЛ=0) и денатурированному (PРАСПЛ=1) состояниям. В зоне перехода экстраполированная (пунктиром) величина PРАСПЛ |экстрап. меняется от 0 до 1 (то есть DPРАСПЛ |экстрап. = 1), а температура меняется на DT. Значит (dPРАСПЛ/dT)|T=To = DPРАСПЛ |экстрап./DT = 1/DТ, — то есть DТ определяется (см. Рис.16-3) зоной самого крутого подъема величины PРАСПЛ — или, в общем виде, шириной зоны резкого изменения любого экспериментального параметра, определяющегося вероятностью расплавленного состояния (например, шириной зоны резкого изменения спиральности белковой цепи).
В окончательной форме, связь эффективной теплоты плавления "плавящейся единицы" (DE) с шириной (DT) и температурой (T0) перехода имеет вид 1/DT = (1/4) DE/kT02, или
DE = 4kT02/DT. | (16.4) |
Рассчитанная таким образом — только из наблюдаемой формы перехода — теплота DE денатурации "единицы плавления" сравнивается с калориметрической теплотой плавления целого белка (рассчитываемой как DH/N, где DH количество тепла, поглощенного всеми имеющимися в калориметре N молекулами белка). Если
DE = DH/N | (16.5) |
— значит, плавление целого белка является переходом типа "все или ничего". Это и есть "критерий Вант-Гоффа".
Если DE < DH/N (т. е. если ширина перехода DT больше, чем того требует критерий Вант-Гоффа), — то "единица плавления" меньше, чем весь белок, т. е. он плавится по частям. Если DE > DH/N, — то "единица плавления" больше белка, т. е. плавится как целое не одна молекула белка, а какой-то белковый агрегат.
Разрушение структуры белка при повышении температуры — плавление — выглядит естественно; однако существует и "холодовая" денатурация белка при аномальном понижении температуры (Рис.16-4). Правда, для многих белков такая денатурация не наблюдается, так как вода в приборе успевает замерзнуть раньше...
Рис.16-4. Обратимая "холодовая" денатурация белка (апомиоглобина) при аномальном понижении температуры (нижняя кривая; стрелка показывает, что температура в ходе опыта понижается). Верхняя кривая показывает ренатурацию белка (левый пик) при обратном повышении температуры и его дальнейшую денатурацию (правый пик). Кривые относятся к избыточной теплоемкости раствора белка по сравнению с теплоемкостью растворителя; они не отнормированы на количество белка в растворе. Картинка взята из Yu. V.Griko, P. L.Privalov, S. Yu. Venyaminov & V. P.Kutyshenko, J. Mol. Biol. (1988) 202:127-138. |
|
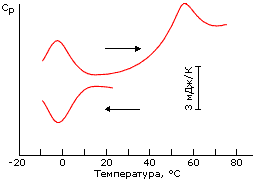
Трудами лаборатории Привалова, где "холодовая" денатурация была впервые обнаружена, было показано также, что она — как и обычная тепловая денатурация — происходит как переход типа "все или ничего".
Причина "холодовой" денатурации кроется в том, что гидрофобные взаимодействия, которые держат белок компактным, сильно растут с температурой — то есть сильно падают с ее понижением. Рост гидрофобных эффектов — и, в частности, их энергии — с температурой проявляется в относительно высокой теплоемкости денатурированного, менее компактного состояния белка (см. DCpd на Рис.16-2). В результате теплота плавления белка сильно растет с температурой — и падает с понижением температуры (Рис.16-2, 16-5а). Падает до того, что может изменить знак при низких температурах (Рис.16-5а)! То есть если обычно более упорядоченная структура (нативный белок) имеет более низкую энергию, чем менее упорядоченная (денатурированный белок) — то при аномально низких температурах (~10оС и ниже) энергия нативного белка может быть выше, чем денатурированного. То же самое, однако, происходит и с энтропией денатурации: она, как и положено энтропии гидрофобного эффекта, тоже падает с падением температуры. В результате стабильность белка (разность свободных энергий его нативной и денатурированных форм) проходит через максимум при "комнатных" или нулевых температурах, — и начинает падать при более значительном охлаждении (Рис.16-5б). При этом иногда стабильность белка падает до того, что при аномально низкой (с точки зрения физиологии: ~0оС и ниже) температуре белок разваливается. Правда, обычно дело до этого не доходит, т. к. вода с белком замерзает, и все процессы крайне замедляются (что позволяет хранить белок на холоду).
Парадоксально, но при холодовой денатурации дело обстоит в точности так, как при кипении воды. Как известно, кипения можно добиться двумя путями — либо подняв температуру, либо понизив внешнее давление. Гидрофобное давление, сжимающее белковую цепь, сильно падает при уменьшении температуры — и белок может "закипеть".
Кстати, действительно "закипеть", — если при повышении температуры белок как бы "плавится", то есть его объем растет не слишком сильно, то при холодовой денатурации белковая цепь совершенно разворачивается и занимаемый ею объем растет многократно.
а | б |
|
|
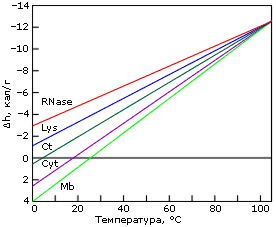
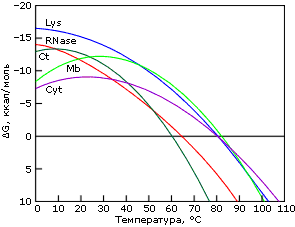
Рис.16-5. Изменение удельной (на грамм белка) разности энергий нативного и денатурированного состояний белка с температурой (а), и изменение свободных энергий в расчете на моль белковых молекул (б). Картинки взяты из P. L.Privalov & N. N.Khechinashvili, J. Mol. Biol. (1974) 86:665-684.
Это естественно подводит нас к вопросу о том, как выглядит денатурированный белок.
Многочисленные термодинамические опыты показывали, что в области денатурированного состояния белковой молекулы в ней нет никаких кооперативных структурных превращений. Поэтому первоначально полагали, что денатурированный белок всегда является очень рыхлым клубком (каковым он, действительно, является в хороших растворителях типа концентрированных растворов мочевины).
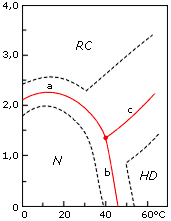
Однако столь же многочисленные структурные исследования денатурированных белков упорно свидетельствовали о каких-то крупных перестройках в рамках денатурированного состояния, о каких-то "промежуточных" структурных состояниях между полностью развернутой (клубковой) и нативной формами молекулы (Рис.16-6).
Вообще, экспериментальные данные о состоянии белковой молекулы после денатурации были очень противоречивы. В одних случаях белки казались полностью "развернутыми" и бесстуктурными, в других — довольно структурированными и/или компактными, а для их полного разворачивания нужны были сильные растворители.
Рис.16-6. Фазовая диаграмма (Танфорд, 1968) конформационных состояний лизоцима при рН 1.7 в растворе гуанидингидрохлорида при различных температурах. N — нативное состояние, RC — клубок ("random coil"), HD — температурно-денатурированное состояние. Жирная линия показывает середину перехода, пунктир — зону перехода (по Танфорду: предположительно, от соотношения 9:1 в пользу одного состояния до 1:9 в пользу другого). Картинка взята из C. Tanford, Adv. Prot. Chem. (1968) 23:121-218. |
|

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
