
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
При преимущественном связывании переходного состояния ведущую роль может играть как связывание его деформированной электронной системы (вспомним "оксианионовую дыру" сериновых протеаз), так и преимущественное связывание деформированной — опять-таки в переходном состоянии — конформации всей молекулы.
Последнее особенно ярко проявляется в каталитической функции искусственных "абзимов" (antybody enzyme) — антител, отобранных так, чтобы, связывая переходное состояние субстрата, они катализировали бы его химическое превращение (Рис.20-12). Некоторые абзимы способны направить на снижение активационного барьера почти всю энергию, выгаданную при связывании субстрата — то есть почти вся она прилагается к деформации именно обрабатываемой химической связи. И хотя имеющиеся абзимы — не очень мощные ферменты (они ускоряют спонтанную реакцию максимум на 5 порядков — а не на 10-15, как естественные ферменты — ведь в них еще не умеют встраивать доноры и акцепторы электронов), — возможность их создания подтверждает важность преимущественного связывания именно переходного состояния.
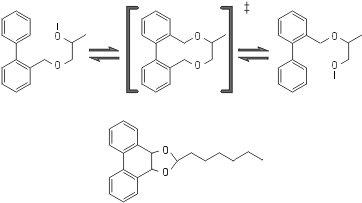
Рис.20-12. Принципиальная схема химического превращения, катализируемого абзимом — искусственно выработанным антителом к переходному состоянию реакции, которая спонтанно идет медленно. В переходном (‡) состоянии изображенной реакции изомеризации имеется — в отличие и от начального, и от конечного ее состояний — плоская система из трех колец. Для выработки антител к переходному состоянию такой формы используется тоже включающая три кольца, но стабильная молекула, изображенная внизу.
Теория катализа за счет преимущественного связывания переходных состояний была обоснована Холденом и Полингом еще в 30-40-х годах. В последние годы переходные состояния для ряда энзиматических реакций были "прощупаны" методами белковой инженерии, т. е. путем целенаправленной замены аминокислот активного центра фермента. При этом удалось выяснить (и увидеть, при помощи рентгена), какие из аминокислотных остатков фермента отвечают за слипание разных компонентов, потребных для создания активированного комплекса, и какие — за преимущественное связывание именно и только переходного состояния реакции, дотоле гипотетического, так как его устройство не было видно структурному эксперименту.
Так, в тирозил-тРНК синтетазе, исследованной А. Ферштом., были найдены те остатки (Thr40 и His 45, см. Рис.20-13), что связываются с g-фосфатом АТФ только в перех![]() дном состоянии, — что и отличает это переходное состояние от просто связанных субстратов или продуктов реакции.
дном состоянии, — что и отличает это переходное состояние от просто связанных субстратов или продуктов реакции.
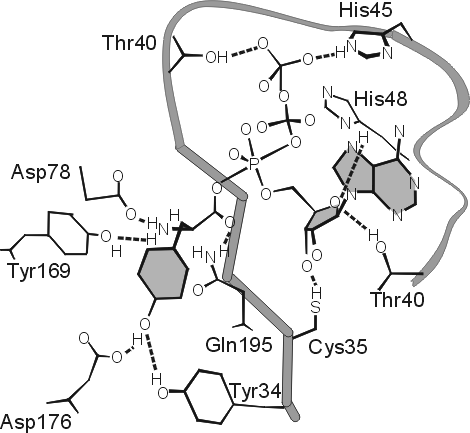
Рис.20-13. Предполагаемое строение переходного состояния в реакции образования тирозил-АМФ из тирозина и АТФ на ферменте тирозил-тРНК синтетазе. В целях наглядности, кольцо тирозина заштриховано, и кольца аденозина — тоже. Атакующий тирозин активированный пяти-координатный a-фосфор (Р) АТФ находится в центре рисунка, а g-фосфат АТФ — в его верхней части. Этот g-фосфат образует водородные связи с Thr 40 и His 45 только в переходном состоянии реакции; при простом, непродуктивном связывании субстратов этих Н-связей нет. Картинка взята из [9].
Перех![]() дное состояние очень нестабильно (по определению: максимально нестабильно!), и прямо увидеть его нельзя. Так что рентген, видящий лишь стабильное расположение субстрата на ферменте, не видит его (субстрата) связывания с Thr40 и His 45 (видимое расстояние слишком велико), между тем как замены Thr40 и His 45 на маленькие аланины снижают активность мутантного белка более чем в миллион раз — хотя эти замены почти совсем не влияют на связывание "мутантом" субстратов реакции. Это свидетельствует, что их (Thr40 и His 45) связывание с g-фосфатом АТФ происходит только в переходном состоянии — при напряженном (пяти-координатном) состоянии g-фосфата, атакованного СО-группой тирозина.
дное состояние очень нестабильно (по определению: максимально нестабильно!), и прямо увидеть его нельзя. Так что рентген, видящий лишь стабильное расположение субстрата на ферменте, не видит его (субстрата) связывания с Thr40 и His 45 (видимое расстояние слишком велико), между тем как замены Thr40 и His 45 на маленькие аланины снижают активность мутантного белка более чем в миллион раз — хотя эти замены почти совсем не влияют на связывание "мутантом" субстратов реакции. Это свидетельствует, что их (Thr40 и His 45) связывание с g-фосфатом АТФ происходит только в переходном состоянии — при напряженном (пяти-координатном) состоянии g-фосфата, атакованного СО-группой тирозина.
Говоря о ферментативной реакции, мы не можем упускать из виду также ее очень высокую специфичность.
Так, сериновые протеазы щепят полипептид только после определенных аминокислот: химотрипсин — после ароматических, трипсин — после положительно заряженных, эластаза — только после самых маленьких. Этот эффект [он называется опознаванием субстрата по принципу "ключ (субстрат) — замок (фермент)"] достигается четкой структурой той специфической части субстрат-связывающего "кармана" (Рис.20-14), куда должна улечься последняя (как на трипсине) или предпоследняя (как на папаине) перед точкой щепления боковая группа пептида.
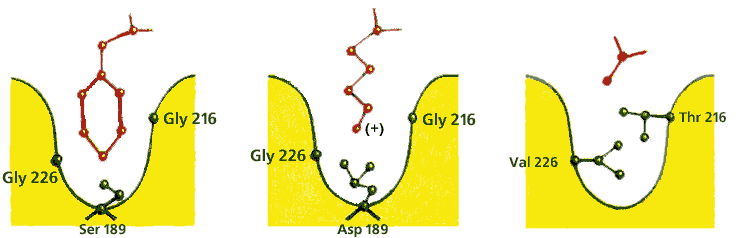
Рис.20-14. Устройство специфического сайта субстрат-связывающего кармана в разных сериновых протеазах. Картинка взята из [5].
Протеазы еще могут позволить себе изредка ошибаться в выборе точки щепления.
Но есть белки, от которых требуется колоссальная точность работы. Это, например, аминоацил-тРНК синтетазы: они "заряжают" тРНК нужными аминокислотами, и от точности их работы зависит точность синтеза белков. Поэтому они не могут позволить себе ошибаться чаще, чем однажды на много тысяч синтезов аминоацил-тРНК, — а опознавание аминокислоты специфическим субстрат-связывающим карманом допускает порядка 1% ошибок. Для устранения этих ошибок используется эффект "двойного сита" (Рис.20-15). Этот эффект обеспечивается специальной конструкцией фермента.
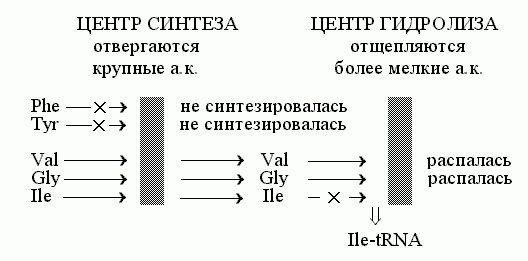
Рис.20-15. Схема "двойного сита" для механизма редактирования работы изолейцил-тРНК синтетазы
У фермента аминоацил-тРНК синтетазы два активных центра: центр синтеза и центр гидролиза. Причем гидролиз аминоацил-тРНК на аминоацил-тРНК синтетазе не есть просто реакция, в точности обратная синтезу: обе эти реакции идут с выделением свободных фосфатов, т. е. обе — с понижением свободной энергии.
При этом субстрат-связывающий карман центра гидролиза меньше, чем карман центра синтеза аминоацил-тРНК.
Принцип сита — не пропускать частицы, большие, чем ячейки сита.
На первом сите — при синтезе аминоацил-тРНК — тРНК заряжается "правильной" аминокислотой и некоторым числом "неправильных", более мелких (более крупные аминокислоты отвергаются все, а большая часть "неправильных" мелких все же отвергается по причине различий в гидрофобности/гидрофильности, но отбор по гидрофобности/ гидрофильности не столь жесток, как отбор по возможности втиснуться в карман данного размера). Итак, на выходе этапа синтеза, — много аминоацил-тРНК с "правильной" аминокислотой и некоторое количество аминоацил-тРНК с "неправильными" аминокислотами, — но только с теми, что меньше, чем "правильная": более крупные аминокислоты отвергаются "карманом" синтетического центра, этим первым "ситом".
На втором активном центре, на втором "сите" — у него "ячейка" (субстрат-связывающий карман) поменьше — идет гидролиз образовавшихся аминоацил-тРНК, — но только тех, где аминокислота меньше, чем "правильная". То есть "правильная" аминокислота отвергается карманом гидролизного центра, этим "вторым ситом", а все остальные аминоацил-тРНК гидролизуются и распадаются.
В сумме, два эти сита и обеспечивают выход только "правильно заряженных" аминоацил-тРНК.
В завершение рассказа о специфичности ферментов, стоит остановиться на двух прагматических аспектах. Во-первых, сейчас масса людей занята изучением субстрат-связывающих карманов и подбором — с открытыми глазами, не на ощупь — связывающихся с ними ингибиторов. Особый интерес, по понятным причинам, вызывают протеазы и другие белки вируса СПИДа. Во-вторых, — большое применение находят себе точечные мутации в районе активного (и прежде всего субстрат-связывающего) центра: они позволяют модифицировать "природную" субстратную специфичность (например, менять специфичность сериновых протеаз) и даже создавать новые, искусственные специфичности для нужд промышленности и медицины. Значительный интерес также вызывают попытки — пока это лишь попытки — создания (на "твердом фундаменте" белковой глобулы) "нового" активного центра, — с новой, "привитой" функцией, — путем направленных точечных замен аминокислотных остатков исходного белка.
Изучение ферментативной деятельности белка показывает (Рис.20-9), что в нее вовлечена совсем небольшая часть белковой глобулы, — в то время как остальная ее часть служит лишь как бы твердым каркасом, обеспечивающим "правильное" строение закрепленного на нем активного центра.
Поэтому нас не должно удивлять, что белки с совсем разной первичной структурой, и даже с совсем разной и пространственной структурой могут иметь одинаковые или очень сходные биохимические функции.
Классическим примером снова являются сериновые протеазы. Есть два класса таких протеаз — типа трипсина и типа субтилизина. Они совсем не похожи ни по аминокислотной последовательности, ни по общей форме (Рис.20-16), и даже принадлежат к разным структурным классам (трипсин — двухдоменный b-белок, субтилизин — однодоменный a/b-белок). Они имеют одинаковую конфигурацию лишь ключевых аминокислотных остатков в каталитическом центре (Рис.20-17) — причем не всех остатков целиком, а лишь их функционально-важных "кончиков" — но не имеют одинаковой конфигурации остатков даже в субстрат-связывающем кармане.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
