
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Разобраться в этой противоречивой картине удалось только, исследуя структуры денатурированных белков целым букетом методов. В этот букет входили и измерения вязкости (дающие объем молекул белка), и спектры КД в дальнем ультрафиолете (отражающие вторичную структуру белка), и спектры КД в ближнем ультрафиолете (отражающие асимметрию, т. е. упорядоченность окружения ароматических боковых групп), — а также и инфракрасная спектроскопия, и ЯМР, и исследование активности белков, и многое другое. Весь этот арсенал и был применен к исследованию денатурации белков группой , куда и я имел честь принадлежать в качестве теоретика.
Оказалось, что, кроме активности белка, есть только две вещи, которые всегда резко меняются при денатурации. Это — (1) упорядоченность окружения боковых групп белка, наблюдаемая как по КД в ближнем ультрафиолете (Рис.16-7а), так и при помощи ЯМР, и (2) фиксированность глобулярной структуры, наблюдаемая — посредством ЯМР — по скорости обмена водородов (Н) полярных групп белка на дейтерий (D) окружающей воды, а также по ускорению протеолиза белковой цепи.
a | б |
|
|
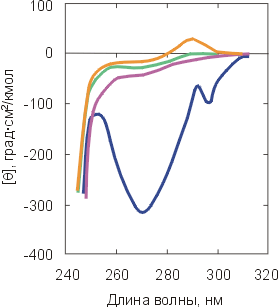
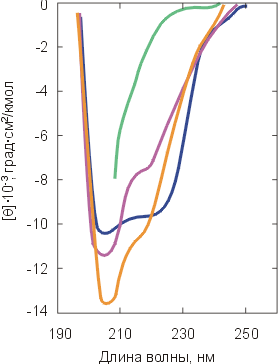
Рис.16-7. Спектры КД в ближнем (а) и дальнем (б) ультрафиолете для нативного a-лактальбумина (![]() ) и для него же, денатурированного теплом (
) и для него же, денатурированного теплом (![]() ), кислотой (
), кислотой (![]() ) и сильным денатурантом (
) и сильным денатурантом (![]() ). Спектры КД в ближнем УФ всегда сильно меняются при денатурации. Спектры КД в дальнем УФ сильно меняются только при сильном разворачивании белка (см. кривую
). Спектры КД в ближнем УФ всегда сильно меняются при денатурации. Спектры КД в дальнем УФ сильно меняются только при сильном разворачивании белка (см. кривую ![]() ). Картинки взяты из D. A.Dolgikh, L. V.Abaturov, I. A.Bolotina, E. V.Brazhnikov, V. E.Bychkova, V. N.Bushuev, R. I.Gilmansin, Yu. O.Lebedev, G. V.Semisontov, E. I.Tiktopulo & O. B.Ptitsyn, Eur. Biophys. J. (1985) 13:109-121.
). Картинки взяты из D. A.Dolgikh, L. V.Abaturov, I. A.Bolotina, E. V.Brazhnikov, V. E.Bychkova, V. N.Bushuev, R. I.Gilmansin, Yu. O.Lebedev, G. V.Semisontov, E. I.Tiktopulo & O. B.Ptitsyn, Eur. Biophys. J. (1985) 13:109-121.
Степень же упорядоченности главной цепи белка (ее вторичная структура), размеры глобулы и даже плотность ее гидрофобного ядра могут как практически сохраняться, так и меняться очень сильно (Рис.16-7б).
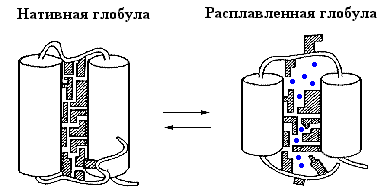
В ходе этих исследований был обнаружен весьма универсальный интермедиат разворачивания (и сворачивания) белков, позже названный "расплавленной глобулой" (Рис.16-8, 16-9). Важнейшие свойства этого интермедиата суммируются Таблицей 16/1.
Рис.16-8. Относительное изменение спектров КД a-лактальбумина в ближнем ( |
|
| Рис.16-9. Схематическая модель нативной и расплавленной глобулы в молекуле белка. Для простоты, на рисунке показаны только две a-спирали, соединенные петлей. Каждая вторичная структура покрыта множеством боковых групп; прошитая водородными связями, вторичная структура стабильна (так что свободная энергия внутримолекулярных водородных связей не меняется), пока проникающий внутрь растворитель не "растворит" всю глобулу. Вода обычно не способна это сделать без добавления денатуранта. Входящие в гидрофобное ядро белка боковые группы заштрихованы. В расплавленной глобуле они обретают свободу движений, выигрывают энтропию, но теряют энергию плотного контакта. Так в глобуле возникают пoры, куда вода ( |
Для очень многих (хотя и не для всех) белков "расплавленная глобула" появляется (путем перехода "все-или-ничего") при умеренном денатурирующем воздействии на нативный белок и исчезает (превращается в неупорядоченный клубок) лишь с добавлением концентрированного денатуранта. "Расплавленная глобула" обычно не плавится при дальнейшем нагреве, но ее разворачивание в клубок сильным растворителем выглядит как кооперативный S-образный переход.
Некоторые нативные белки, однако, разворачиваются прямо в клубок, минуя стадию расплавленной глобулы.
Противоречивость результатов структурных и термодинамических исследований денатурации белков, а также физическая природа нового фазового состояния белков — состояния "расплавленной глобулы" — требовали теоретического осмысления.
Для понимания природы состояния расплавленной глобулы важно, что получается из нативного состояния путем кооперативного температурного плавления — фазового перехода первого рода. Это означает, что оно обладает значительно большей энтальпией и энтропией, чем нативное состояние, то есть что внутримолекулярные взаимодействия в нем резко ослаблены, а подвижность белковой цепи — резко увеличена. Так как большинство внутренних степеней свободы в белке связано с мелкомасштабными флуктуациями структуры, и прежде всего с движениями боковых групп, именно раскрепощение таких флуктуаций может сделать состояние расплавленной глобулы термодинамически выгодным. Однако раскрепощение мелкомасштабных флуктуаций не требует полного разворачивания белковой цепи — достаточно лишь ее небольшого набухания. При этом, однако, в белке резко ослабляется Вандерваальсово притяжение: оно сильно зависит от расстояния и даже небольшого увеличения размеров молекулы достаточно для его значительного ослабления (Рис.16-9).
В общем, здесь все похоже на плавление кристалла: небольшой рост его объема рвет часть Вандерваальсовых взаимодействий в нем и раскрепощает движения молекул.
"Расплавленная глобула" послужила и ключом к пониманию кооперативности денатурации белка. Оказалось, что этот процесс качественно отличается от переходов глобула-клубок в "нормальных" полимерах.
Дело в том, что как эксперименты с гомополимерами (крайне трудные: из-за угрозы агрегации их приходится проводить при предельно низких концентрациях полимера), так и физические теории гомополимеров и статистических гетерополимеров (не "отобранных", как белки, а "нормальных", т. е. "случайных") расходились с тем, что наблюдается в белках.
Они все говорили, что разрушение плотной глобулы в полимерах должно начиниться постепенно, и до самого (или почти до самого) конца идти без резкого скачка плотности молекулы, связанного с переходом одной фазы ("глобула") в другую ("клубок"), что к моменту превращения глобулы в клубок она уже становится очень рыхлой, и что эти две фазы различаются в основном масштабом флуктуаций плотности молекулы (малым — в глобуле, и большим — в клубке), — то есть что этот переход совсем не похож на фазовый переход первого рода — скажем, на испарение. А денатурация белка похожа как раз на плавление или испарение твердого тела.
Несмотря на то, что классическая теория переходов глобула-клубок в гомополимерах не применима к денатурации белков, — я считаю не лишним рассказать о ней — естественно, в самой упрощенной форме. Такой рассказ, с одной стороны, позволит мне позже оттенить то, что существенно именно для белковых, "отобранных" цепей, а не просто для полимеров. С другой стороны — эта теория вполне применима к поведению уже денатурированных белковых молекул. И, наконец, я считаю, что знание базовых физических моделей необходимо для общей культуры.
Я рекомендую вам "поверять слова формулами" при чтении дальнейшего текста. Конечно, проще читать только слова, но они часто допускают неоднозначное толкование; так что, поверяя слова формулами, — вы сами проверите свое понимание.
Итак, я рассмотрю простейшую "полимерную" модель, — много (N>>1) одинаковых и связанных в цепь мономеров ("бусинок"). И я сравню ее с простейшей моделью, используемой со времен Ван дер Ваальса (XIX век!) для описания переходов газ-жидкость — с "облаком мономеров", т. е. с совокупностью из N таких же, но не связанных в цепь мономеров (Рис.16-10).
Облако мономеров | Полимерная цепь | ||
|
|
|
|
|
| ||
|
| ||
T1 < T2 < T3 < T4 | |||
|
|
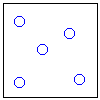
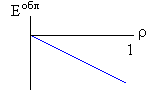
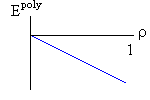
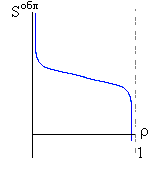
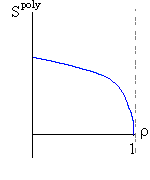
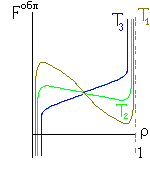
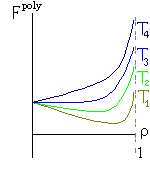
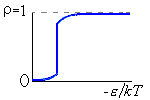
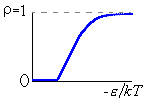
Рис.16-10. Качественное сравнение поведения зависимости энергии DЕ, энтропии DS и свободной энергии DF=DЕ-TDS добавления одного мономера от плотности r для двух систем: "облако мономеров" и "полимерная цепь". Графики для энергий приведены для притягивающихся (e<0) мономеров. Зависимости DF от r показаны для разных температур Т. На дополнительных графиках (внизу) показана качественная зависимость плотности r обеих систем от энергии взаимодействия мономеров, выраженной в единицах kT; графики относятся к невысокому внешнему давлению [при очень высоком, "надкритическом" давлении разреженных (с r<0.5) состояний не существует].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
