
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
И последнее на сегодня. Имея графики зависимости и энтропии S, и энергии Е от плотности глобулы r, мы можем построить график зависимости S от E и, далее, нарисовать характерный вид энергетического спектра белка (Рис.16-15). Основной чертой этого спектра является отрыв энергии самой стабильной, т. е. нативной структуры белка от энергий, где начинается спектр подавляющего большинства остальных структур белковой цепи. Именно этот отрыв (или, как говорят, "энергетическая щель между самой стабильной структурой и ее конкурентами") и заставляет белок плавиться путем перехода "все-или-ничего" — а до того "терпеть", не меняясь, изменение внешних условий.
а | б |
|
|
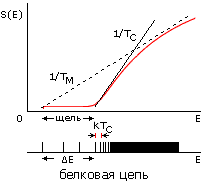
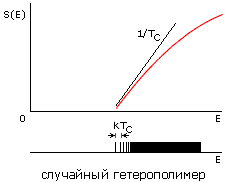
Рис.16-15. Зависимости энтропии S от энергии Е для белковой цепи (а) и для цепи случайного гетерополимера (б). Показаны касательные к нижней части кривых S(E), определяющие температуру TC стеклования случайного полимера (она же фигурирует в статистике белковых структур) и температуру TМ плавления белка (наклон второй касательной несколько меньше, т. е. TМнесколько выше TС). Внизу показаны характерные формы энергетических спектров этих молекул. Каждая линия спектра (здесь показаны лишь немногие из этих линий) соответствует одной конформации цепи. В правой части обоих спектров этих линий очень много — и они сливаются. Основной чертой спектра белка — "отобранного" гетерополимера — является энергетическая щель (ширины DЕ >> kTC) между основной, самой стабильной укладкой цепи и другими, не похожими на нее укладками. Немногие структуры, чьи энергии приходятся на эту щель, представляют собой более или менее "разболтанные" или "недосвернутые" варианты основной укладки цепи.
Теорфизические исследования разнообразных моделей гетерополимерных цепей показали, что в случайных гетерополимерах такая щель не наблюдается. Точнее — у случайных цепей она обычно очень мала, так что такие цепи замерзают (при температуре TC, о которой — чуть ниже) не скачком, как кристалл или белок, а постепенно, как стекло.
Значит, необходимая для фазового плавления белка достаточно большая (с шириной DЕ >> kTC), энергетическая щель между самой стабильной структурой цепи и ее конкурентами возникает не в любой — "случайной" — аминокислотной последовательности, а создается отбором "белковоподобных", годных для создания белков последовательностей (в частности — последовательностей, допускающих плотную упаковку цепи в глобулу).
Удалось даже ориентировочно оценить долю "белковоподобных" среди всех случайных цепей:
ДОЛЯ(DЕ) ~ ехр(-DЕ/kTC) . | (16.10) |
Температура TC определяется наклоном кривой S(E) перед самой щелью (Рис.16-15); TC не зависит от величины DЕ, т. е. она одинакова и для случайных, и для "белковоподобных" цепей одинакового аминокислотного состава. И это — та самая температура TC, что фигурирует в белковой статистике, о которой мы говорили на прошлой лекции. Рис.16-15 показывает, что, при не слишком больших DЕ, температура TC лежит чуть ниже температуры плавления белка. А очень большие щели, большие DЕ, по-видимому, не нужны для "белковоподобного" поведения молекулы, так что отбор на них настаивать не должен; и в то же время они маловероятны, согласно формуле 16-10.
Энергетическая щель — фундаментальная вещь в физике белка. Она необходима не только для того, чтобы белок разрушался бы только фазовым переходом, — т. е. чтобы он, не изменяясь, терпел бы (до известного предела) изменение внешних условий, т. е. для надежности работы белка. Скоро вы увидите, что она необходима и для быстрого и безошибочного сворачивания его нативной структуры.
Лекция 17
Сегодня мы посмотрим, как протекает процесс обретения белковой цепью своей нативной структуры во времени, и обсудим кинетику самоорганизации белков. Я буду говорить только о водорастворимых глобулярных белках. Самоорганизация мембранных белков изучена еще сравнительно мало. Самоорганизация фибриллярных белков изучена лучше, но скорее не на физическом, а на биохимическом уровне (она уже обсуждалась на примере образования коллагена).
В живой клетке белок синтезируется на рибосоме. Биохимический синтез белковой цепи занимает порядка минуты, да и все изготовление "готового", свернутого белка, занимает примерно столько же — эксперимент не видит разницы (для справки: весь жизненный цикл бактерии может длиться всего пару десятков минут). Поэтому естественно предположить, что сворачивание белка может начинаться еще на рибосоме, еще до окончания ее полного синтеза всей белковой цепи.
К сожалению, достоверные экспериментальные данные об образовании белковой структуры in vivo — в клетке — весьма скудны: очень трудно увидеть структурные превращения растущего белка на фоне всего клеточного супа. Обычно приходится останавливать синтез, выделять "недоделанный" белок, исследовать его отдельно — на все уходит время, десятки минут, в течение которых пространственная структура исследуемой белковой цепи может кардинально измениться...
Ряд такого рода опытов с большими белками показывают, что их первые, N-концевые домены успевают (или, точнее, обладают способностью) свернуться еще до окончания полного синтеза цепи.
Здесь, однако, надо подчеркнуть, что все данные эти относятся к многодоменными белкам; это существенная оговорка, так как "единицей самоорганизации", по-видимому, является не белок в целом, а его отдельный домен.
Об этом свидетельствуют два ряда данных, полученных, правда, не in vivo, а in vitro: во-первых, отдельно взятые домены часто способны к правильной самоорганизации; во-вторых, однодоменный белок, лишенный примерно десятка аминокислот на своем С-конце, к самоорганизации не способен.
Ряд интересных данных по сворачиванию белков получен в бесклеточной белок-синтезирующей системе (т. е. не совсем in vivo, но и не совсем in vitro). Такая система состоит из рибосом, транспортных и информационных РНК и других факторов, необходимых для матричного синтеза белковой цепи.
К сожалению, и здесь очень трудно увидеть строение растущего белка на фоне громадной рибосомы. Строение, действительно, увидеть трудно. Но, если повезет, можно увидеть активность вновь синтезированного белка.
Счастливой находкой здесь оказалась люцифераза: этот белок, приняв свою нативную конформацию, катализирует реакцию с испусканием света. Так что появление нативной люциферазы легко увидеть — ведь больше в клетке ничего не светится. Исследуя биосинтез и сворачивание этого белка, — , , и в нашем институте показали, что первый активный белок появляется через 10 мин. после включения его биосинтеза, а выключение, блокирование биосинтеза немедленно прекращает выход активного белка (Рис.17-1). Это значит, что цепей уже синтезированных, но еще не свернувшихся, практически нет — то есть показывает, что сворачивание этого большого, содержащего свыше 500 остатков в цепи белка идет либо котрансляционно, либо почти мгновенно после конца биосинтеза.
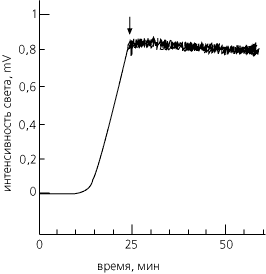
Рис.17-1. Свечение нативной люциферазы, синтезируемой в бесклеточной системе. Время "0" — включение биосинтеза люциферазы. Время выключения биосинтеза указано стрелкой. Сразу после этого свечение прекращает свой рост, т. е. выход нового активного белка больше не наблюдается. Картинка взята из V. A.Kolb, E. V.Makeev & A. S.Spirin, EMBO J. (1994) 13:3631-3637.
Однако для выяснения не только результата, но и хода процесса самоорганизации приходится исследовать самоорганизацию белка "совсем in vitro" (без всяких рибосом, шаперонов и т. д.), — то есть при ренатурации белка в растворе.
Глобулярный белок способен к спонтанной самоорганизации in vitro (ренатурации), если после биосинтеза он не подвергся сильной химической модификации. В этом случае его архитектура, "мягко" (без разрыва цепи) разрушенная температурой, растворителем и т. д., — спонтанно восстанавливается при "нормализации" среды. Правда, эффективная ренатурация нуждается в тщательном подборе экспериментальных условий — иначе ей может воспрепятствовать агрегация белка.
Явление спонтанной самоорганизации белков было открыто в группе Анфинсена в 1961 г. на бычьей рибонуклеазе А. Это открытие, впоследствии подтвержденное на множестве других белков, а также возможность чисто химического синтеза белковой цепи, спонтанно сворачивающейся далее в активный белок (опыты Меррифилда и др.), позволяет — в первом приближении — отделить изучение структурообразования белка от изучения биосинтеза белковой цепи.
Самоорганизация белка in vitro может рассматриваться как самый простой (и поэтому для меня, физика, наиболее интересный) случай "чистой" самоорганизации, когда белку не помогает никто.
Вообще, самоорганизация пространственных структур белков (а также РНК) — уникальное физическое явление, не имеющее аналогов в "неживой" природе. Эта самоорганизация напоминает образование кристаллов, — но кристаллов, во-первых, не имеющих периодической пространственной структуры; во-вторых, чрезвычайно сложно устроенных; и, наконец, очень маленьких.
Самоорганизация белковых структур относится, с физической точки зрения, к классу явлений "возникновение порядка из порядка" (по классификации Пригожина): трехмерный "апериодический кристалл" (говоря словами Шредингера) структуры белка порождается заранее фиксированным порядком звеньев в его цепи. Видимо, следует сразу отметить, что самоорганизация трехмерной структуры белков (и РНК, и кристаллов вообще) возникает из стремления молекул к термодинамическому равновесию — и тем принципиально отличается от той самоорганизации типа "порядок из беспорядка" (будь то самоорганизация в осциллирующей химической реакции Белоусова-Жаботинского или самоорганизация в экологической системе "хищник-жертва"), которую обычно имеют в виду, говоря о самоорганизации в неравновесных, работающих на протоке энергии системах.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
