
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Однако прежде, чем перейти к рассказу о самоорганизации белка in vitro, к рассказу о физике этого спонтанного процесса, я хочу кратко упомянуть ту машинерию, которая используется клеткой для повышения эффективности самоорганизации белка.
Рибосома выдает белковую цепь постепенно и не вполне равномерно — есть паузы, приостановки биосинтеза цепи; предполагается, что соответствие пауз границам структурных доменов способствует их спокойному, без вмешательства извне, созреванию.
Далее. В клетке белковая цепь сворачивается под опекой специальных белков — шаперонов. "Малые" шапероны типа hsp70 (heat shock protein, 70 килодальтон) связываются с белком, предохраняя его от агрегации, а потом сбрасываются (на что расходуется АТФ). "Большие" шапероны типа GroEL и GroES или TriC работают в основном с многодоменными белками, и особенно — с белками, чьи домены составлены из отдаленных кусков цепи. Эти шапероны образуют как бы пробирку (диаметром в несколько нанометров — и с крышечкой!), куда поступает новосинтезированный (и предварительно облепленный шаперонами типа hsp70 и/или hsp40) белок или его домен. Эта "пробирка" (иногда ее называют "ячейкой Анфинсена") защищает новорожденный белок и от агрегации, и от действия концентрированного клеточного "супа". При этом пробирка "трясется" и время от времени активно открывается (на что расходуется АТФ) и закрывается, — но отпускает она белок только тогда, когда он уже свернется и перестанет липнуть к пробирке.
Кроме того, самоорганизация белков может ускоряться некоторыми ферментами типа пролил-изомеразы (катализирующей медленно само по себе идущее превращение trans пролинов в cis и обратно) или дисульфид-изомеразы (катализирующей сшивание и расшивание SS связей).
Однако опыт показывает, что работа всей этой клеточной машинерии может быть заменена подбором соответствующих внешних условий (малой концентрации белка, нужного окислительно-восстановительного потенциала). Эта замена не меняет результат сворачивания: уж если белок не выпал в осадок, а свернулся in vitro — то он свернулся в ту же структуру, что и in vivo. Правда, часто это займет больше (а иногда — и меньше!) времени, чем самоорганизация in vivo, — но результат будет тот же.
Все это показывает, что вся необходимая для построения трехмерной структуры белка информация содержится в химической последовательности аминокислот в его цепи.
Похоже, что — помимо самого синтеза белковой цепи — биосинтетический аппарат клетки (рибосома + шапероны + ...) служит лишь чем-то вроде инкубатора для сворачивания пространственной структуры белка: этот "инкубатор" не определяет структуру белка, но он создает условия для ее созревания, — так же, как обычный инкубатор обеспечивает выведение птенца, не предопределяя, кто выведется — цыпленок или утенок.
Загадочность самоорганизации белков (и РНК) суммируется "парадоксом Левинталя". Загадка состоит вот в чем. У белковой цепи есть бездна возможных конформаций (каждый аминокислотный остаток имеет около 10 возможных конформаций, то есть цепь из 100 остатков — порядка 10100 возможных конформаций). Так что белок должен искать "свою" пространственную структуру среди порядка 10100 возможных. И так как переход из одной конформации в другую занимает ~10-13 секунды как минимум, перебор всех 10100 структур должен был бы занять порядка 1080 лет, на фоне которых время жизни нашей Вселенной — 1010 лет — величина бесконечно малая... Вопрос: как белок может "найти" свою структуру за минуты?
Парадокс же заключается в следующем. С одной стороны, нативная пространственная структура по всем тестам ведет себя как самая стабильная из всех структур цепи: белковая цепь попадает в нее при разных кинетических процессах [и при сворачивании на рибосоме в процессе биосинтеза, и после секреции сквозь мембрану, и при сворачивании в пробирке (ренатурации), — чем бы и как бы она ни была в этой пробирке развернута]. С другой стороны, нет никаких гарантий, что эта структура — самая стабильная из всех возможных: у белковой цепи просто нет времени на то, чтобы убедиться в этом!
Как же белок выбирает свою нативную структуру среди бесчисленного множества возможных? — спросил Левинталь, и ответил: — По-видимому, самоорганизующийся белок следует по какому-то специальному "пути сворачивания", и та структура, где этот путь заканчивается, и является его нативной структурой. Иными словами, Левинталь предположил, что нативная структура белка определяется не стабильностью, не термодинамикой, а кинетикой, т. е. она соответствует не глобальному, а просто быстро достижимому минимуму свободной энергии цепи.
Вопрос о том, что именно — кинетика или термодинамика — определяет укладку белковой цепи — отнюдь не чисто умозрительный. Он постоянно возникает на пути решения конкретных задач физики белка, — идет ли речь о предсказании структуры белка по его аминокислотной последовательности (надо знать, что предсказывать: самую стабильную или самую быстро сворачивающуюся его структуру), или о дизайне новых, не встречающихся в природе белков (надо знать, что делать: максимально усиливать стабильность желаемой структуры или пролагать максимально быстрый путь к ней).
Обсуждение механизма сворачивания белков началось сразу же после расшифровки первых трехмерных структур и открытия явления самоорганизации. Первой, видимо, была гипотеза Филлипса, согласно которой на N-конце растущей цепи (с которого начинается биосинтез белка) возникает зародыш структуры, и остальная цепь наматывается на него. В той или иной форме такая точка зрения присутствует в ряде работ и по сей день. Однако она не подтвердилась. В изящных работах Гольденберга и Крейтона было показано, что N-конец цепи не играет решающей роли в самоорганизации in vitro. Замкнутая в кольцо цепь небольшого белка, ингибитора трипсина, сохраняет способность к самоорганизации; и даже если разрезать это кольцо так, что новым N-концом окажется бывшая середина цепи — самоорганизация ведет к прежней пространственной структуре.
В поисках пути к решению проблемы самоорганизации белков, в 1973 году предложил концепцию стадийного сворачивания белка (Рис.17-2). В рамках этой гипотезы, получившей позже название "framework model" ("каркасной модели"), и проходило, в значительной мере, дальнейшее обсуждение проблемы самоорганизации белка в мировой научной литературе.
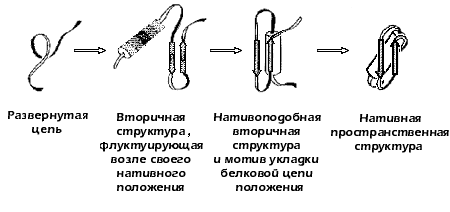
Рис.17-2. Стадийная модель сворачивания белка по (1973). Выделены вторичные структуры — a-спирали (цилиндры) и b-участки (стрелки).
Эта модель постулировала последовательное вовлечение различных взаимодействий в формирование структуры белка и подчеркивала важность образования зародышевых a-спиралей и b-шпилек на ранних стадиях сворачивания, слипания этих зародышей в глобулу, грубо напоминающую нативную, и окончательной "доводки" строения глобулы на последнем этапе самоорганизации.
Ключевым камнем этой концепции было то, тогда чисто гипотетическое, а теперь вошедшее в учебники состояние белковой цепи, которое ныне известно как "расплавленная глобула".
Выведенная буквально "на кончике пера", расплавленная белковая глобула была затем экспериментально обнаружена и детально исследована Долгих, Семисотновым, Гильманшиным и др. в лаборатории Птицына в 80-х годах, — первоначально как равновесное состояние "слабоденатурированного" белка (об этом я говорил на прошлой лекции), а потом и как кинетический интермедиат сворачивания белка in vitro.
Так, расплавленная белковая глобула накапливается в процессе ренатурации карбоангидразы В (Рис.17-3). Видно, что "нативные" значения приведенной вязкости (характеризующей объем глобулы) и эллиптичности при 222 нм (характеризующей наличие вторичной структуры) восстанавливаются в процессе ренатурации значительно быстрее, чем эллиптичность при 270 нм (характеризующая упаковку боковых групп) или активность белка. Это говорит о существовании кинетического интермедиата, — причем интермедиата, являющегося расплавленной глобулой: его компактность и вторичная структура близки к таковым для нативного белка, однако он не имеет нативной упаковки боковых групп и эстеразной активности нативного ("твердого") белка.
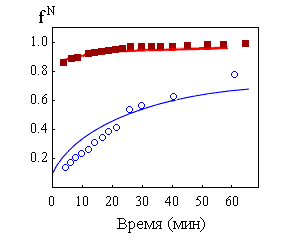
Рис.17-3. Кинетика восстановления "степени нативности" fN в процессе ренатурации карбоангидразы В. Переход белка из полностью развернутого состояния (существовавшего при 5,45 М гуанидингидрохлорида) в нативное (при 0,97 М гуанидингидрохлорида) наблюдается по различным параметрам: приведенной вязкости ![]() , эллиптичности при 222 нм
, эллиптичности при 222 нм ![]() и 270 нм ( _______ ) и энзиматической активности (о о о). Картинка взята из D. A.Dolgikh, A. P.Kolomiets, I. A.Bolotina & O. B.Ptitsyn, FEBS Letters (1984) 164:88-92.
и 270 нм ( _______ ) и энзиматической активности (о о о). Картинка взята из D. A.Dolgikh, A. P.Kolomiets, I. A.Bolotina & O. B.Ptitsyn, FEBS Letters (1984) 164:88-92.
Расплавленная глобула оказывается — в физиологических условиях — ранним промежуточным состоянием на стартующем из клубка пути самоорганизации многих глобулярных белков in vitro. Для образования такого интермедиата достаточно миллисекунд, полное же восстановление свойств нативного белка из 100-300 аминокислотных остатков требует от секунд (для одних белков) до десятков минут (для других).
Необходимо подчеркнуть, что самый медленный (rate-limiting) шаг самоорганизации приходится не на раннюю стадию, не на образование расплавленной глобулы, а на образование из нее плотно упакованной, нативной глобулы (Рис.17-3).
Все описанные выше работы велись, как теперь говорят, в рамках "химической логики", императив которой — "ищи и выделяй промежуточные состояния!". В химии это обычно позволяет понять, как протекает реакция. Но при изучении самоорганизации белков эта логика не сработала: интермедиаты были определены, но ключевые вопросы остались без ответа.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 |
