
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Принципы метода. Допустим, что имеется система, состоящая из точечного источника света и конденсора, преобразующего расходящийся световой пучок в параллельный. На пути этого пучка поставим непрозрачный экран с малым отверстием произвольной формы. Свет будет дифрагмировать на этом отверстии. Поставив выпуклую линзу, проследим, как формируется оптическое изображение светящегося объекта (отверстия) .В фокальной плоскости линзы соберутся параллельные лучи, и будет наблюдаться первичное изображение объекта. Вторичное изображение будет наблюдаться в плоскости, находящейся на определенном расстоянии от фокуса линзы и сопряженной с плоскостью объекта. Дифракционная картина объекта в фокальной плоскости рассчитывается при помощи трансформации Фурье. Задача метода в том, чтобы найти пространственное расположение атомов в молекуле исследуемого вещества по его рентгеновской дифракционной картине. Для вычисления трансформаций Фурье нужно также знать величины амплитуд дифрагированных лучей и их относительные фазы.
Основная трудность метода в том, что из экспериментальных данных не удается измерить фазы дифрагированных лучей.
Рентгеноструктурные исследования кристаллов белков начинаются с определения формы и размеров элементарной ячейки и установления его пространственной группы симметрии, которые входят во все последующие расчеты.
Дифракция рентгеновских лучей ) на кристаллической решетке. Рентгеновские лучи - это электромагнитные волны, которые занимают область спектра между гамма-излучением и коротким ультрафиолетом. В структурных исследованиях используются лучи с длинами волн от 0,5 до 2А. Когда на образец попадает рентгеновский луч, под действием электромагнитной волны луча все атомы исследуемого вещества становятся источниками вторичных рассеянных волн, которые могут интерферировать, т. к рассеяние таких рентгеновских лучей на атомах происходит без изменения длины волны. Дифракционная картина определяется особенностями взаимного расположения атомов в исследуемом веществе. Регулярное и симметричное взаимное положение атомов в кристаллических решетках обуславливает возникновение регулярной и симметричной дифракционной картины, состоящей из дискретных максимумов.
Основной закон дифракции рентгеновских лучей в кристалле определяет зависимость угла отражения рентгеновских лучей q от длины волны используемого излучения l и межплоскостного расстояния d той системы, от которой происходит отражение:
2d sin q = nl, (1.6.34.)
где nl - целое число длин волн.
Это уравнение не дает полного представления о дифракционной картине кристалла, а определяет лишь зависимости между скалярными величинами, а отраженные кристаллом рентгеновские лучи ориентированы определенным образом по отношению к системе координат, связанной с кристаллом и могут быть представлены определенными векторами, на которых строится так называемая обратная решетка.
Рентгенограммы белков. Существуют два способа регистрации рентгеновской дифракционной картины : фотографический и ионизационный.
Чтобы увеличить вероятность попадания узла обратной решетки на поверхность сферы отражений при произвольной ориентации кристаллов низкомолекулярных соединений, используют полихроматическое излучение, а также метод вращения или колебания кристалла.
Обратная решетка кристаллов белков густо усеяна узлами. Наибольшая плотность узлов наблюдается в плоскостях обратного пространства. При таком расположении узлов возможно получить очень богатую и специфическую рентгенограмму неподвижного кристалла на монохроматическом излучении. Плоскости обратной решетки белкового кристалла пересекают сферу отражения по окружностям. Отраженные лучи соответствующие узлам, расположенным на таких окружностях, направлены вдоль образующих конуса с вершиной в центре сферы отражений. Такие конусы отраженных лучей пересекаются с плоской фотопленкой по эллипсам, гиперболам или окружностям. По таким рентгенограммам иногда удается оценить размеры элементарных ячеек, однако они дают слишком мало информации о структуре белкового кристалла.
Более эффективны методы, позволяющие получить на фотопленке неискаженные изображения узлов обратной решетки. Рентгенограммы, полученные такими методами содержат по несколько сотен дифракционных максимумов и легко измерять относительные интенсивности рефлексов.
Основная идея методов получения неискаженных изображений обратной решетки кристалла сводится к следующему. Если заставить кристалл и пленку двигаться так, что во время съемки расстояние между кристаллом и пленкой не будет меняться и плоскость пленки все время будет оставаться параллельной плоскости обратной решетки, можно сохранить принцип подобия, и на пленке получится неискаженное изображение узлов в этой плоскости.
Разные методы фотографирования обратной решетки отличаются один от другого характером движения кристалла.
Для измерения интенсивностей отраженных кристаллом рентгеновских лучей применяются дифрактометры, которые позволяют измерять положения и интенсивности рентгеновских отражений при помощи гейгеровских, пропорциональных или сцинтилляционных счетчиков, которые, двигаясь синхронно с кристаллом, регистрируют последовательно дифракционные максимумы.
Преимущества перед фотографическим методом:
¾ при использовании автоматических дифрактометров сокращается время для измерения интенсивностей рефлексов полного дифракционного поля.
¾ точность измерения, определяемая длительностью регистрации максимумов, может быть доведена до 1-2% (фотометод дает 10-15%).
1.6.21. Спектрофотометрия в области электронных переходов.
Спектры белков. В белках группа, дающая характерную полосу поглощения в окрестности 190 ммк ( амидная группа ), образуется при поликонденсации аминокислот, т. к. она является пептидной связью. Поэтому белок можно представить в виде полимера, состоящего из мономерных звеньев - пептидных групп, связанных между собой через a-углерод, что позволяет полагать об отсутствии между ними существенного обмена электронами и взаимодействие имеет в основном диполь-дипольный характер. в случае белков речь может идти вообще только об изменении поглщения при переходе клубок-спираль, т. к. при гидролизе рвется пептидная связь, обусловливающая поглощение. Пептидная связь является плоской, т. к. p-орбиты делокализованы по трем атомам ( N, C, O ) и связь N-C носит частично двойной характер. В амидной группе имеются три p-орбитали, две из которых заняты в основном состоянии, и одна n-орбиталь - локализована на кислороде.
p®p*- переходы поляризованы в плоскости группы, а n®p*- переход запрещен, но проявляется в области 225 ммк. Также имеет место гипохромный эффект в области 190 ммк, который весьма велик. Он обусловлен взаимодействием длинноволновой p®p*- полосы с более коротковолновой p®p*- полосой.
1.6.22. ИК- спектроскопия.
Происхождение иифракрасных спектров обусловлено главным образом колебаниями атомов в молекуле. Спектр, как правило, состоит из большого числа полос, причем для молекулярного анализа важно то, что часть полос может быть приписана колебаниям отдельных атомов группировок, более или менее сохраняющих свойства в разных молекулах.
Спектр нормальных колебаний молекулы занимает область от 4000 до 50 см -1. Спектр органических соединений момно разбить на отдельные интервалы:
4000 - 2000 см -1 ¾ валентные ( с изменением длин связей при колебании ) колебания связей типа XH, YH 2 (X, Y = C, O, N и др.)
2000 - 1300 см -1 ¾ колебания, обусловленные изменениями длин связей, образуемых относительно тяжелыми, по сравнению с водородом или дейтерием, атомами C, N, O с кратностями связей больше единицы.
1300 - 700 см -1 ¾ скелетные колебания, в которых принимают участие все одинарные связи С - С, С - N, C - O, а также углы типа CCH, CNH и др.
< 700см -1 ¾ деформационные колебания, которые обусловлены изменениями углов, образованных связями типа XY.
Особенностью поглощения белков в твердом состоянии является наличие сильных полос в области 3000 – 3500см -1 и около 1600 – 1400 см -1. Эта особенность обусловлена ионным диполярным состоянием молекулы: в высокочастотной области - валентными колебаниями NH2+ или NH3+ , а полосы 1600 и 1400 см -1 соответствуют двум основным колебаниям карбоксильной группы - COO -. По мере удлинения полипептидной цепи проявление концевых групп становится менее заметным, зато отчетливо наблюдается сильное поглощение пептидной (амидной ) группы. Валентные колебания NH - связи дают сильную полосу около 3300см -1. В области ниже 2000см -1 выделяются несколько сильных полос, обозначаемых Амид-1 (около 1650 см -1 ), Амид-2 (около 1550 см -1 ) и Амид-3 ( около 1200-1300 см -1 ), обусловленных колебаниями в плоскости, и широкая полоса поглощения неплоских деформационных колебаний NH ( Амид-5 ).
Другим характерным признаком амидных колебаний могут служить сдвиги частот в сильно разбавленных растворах полярных растворителей. Этот эффект обусловлен разрывом водородной связи
N - H--- O = C. Наблюдаемые изменения нетрудно объяснить, если представить пептидную группу в виде шариков с пружинками : при разрыве водородной связи силовые коэффициенты связей NH и CO увеличиваются, т. к. уменьшаются длины этих связей , т. е. a 0 в формуле
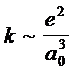 , (1.6.35.)
, (1.6.35.)
тогда как силовые коэффициенты углов OC и CNH уменьшаются. Отсюда следует, что валентное колебание NH в пептидной группе без водородной связи будет иметь более высокую частоту( около 3300см - 1). Точно также и полоса Амид-1 , обусловленная колебанием, при котором сильно изменяется длина связи CO, будет иметь соответственно более высокую частоту ( на 40-50 см -1 ). Частоты деформационных колебаний Амид-2, Амид-3 и Амид-5 сдвигаются при разрыве водородной связи в сторону низких частот. Величина сдвига частоты характеризует силу водородной связи. Наибольший сдвиг имеют валентное и неплоское деформационное колебания группы NH. Сильно изменяются также ширина полос и их интенсивность: при наличии водородной связи полосы поглощения шире и приблизительно на порядок интенсивнее. В большинстве белков полоса валентных колебаний NH - группы имеет частоту максимума в интервале 3280 - 3310 см -1; исключение составляют белки группы коллагена ( 3330 см -1 ). Такая большая стабильность частоты объясняется тем, что основные параметры пептидной водородной связи (главным образом расстояние N --- О ) во всех этих структурах остаются приблизительно постоянными. Частота валентного колебания остается практически неизменной даже тогда, когда белок имеет беспорядочную конформацию цепей, но при этом ширина полосы становится заметно больше.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 |
