
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Расчеты нормальных колебаний пептидной группы показали, что из всех плоских колебаний валентное колебание NH ( n 0 ), полосы Амид-1 и -2 (n1 и n2 ) наиболее локализованы и могут быть описаны из положений равновесия лишь шести атомов самой пептидной группы. Поляризация колебания n1 приблизительно совпадает с направлением связи CO, а для n2 она направлена почти перпендикулярно связи NH в соответствии с направлением поворота этой связи при колебании. Это объясняется тем, что интенсивность и поляризация колебаний пептидной группы в самом грубом приближении могут быть получены из рассмотрения колебаний только тех связей, которые имеют наибольший дипольный момент и характеризуются наибольшим изменением дипольного момента при растяжении связи. Такими связями в пептидной группе являются связи CO и NH. Их дипольные моменты должны быть равны около 2,3 и 1,3 D, если пептидная группа имеет кетонное строение ; на самом деле они еще больше. Все остальные связи имеют дипольные моменты, близкие к нулю, и, следовательно, в этом приближении слабо проявляют себя в интенсивности и поляризации полос поглощения. Благодаря этим сведениям стало возможным успешное изучение структуры полипептидов и фибриллярных белков методом ИК- спектроскопии.
Для примера можно взять картину колебания Амид-1 антипараллельной кристаллической упаковки вытянутых полипептидных цепей. В элементарной ячейке находятся четыре пептидные группы, соединенные между собой попано посредством химических или водородных связей. Согласно общей теории колебаний, в этом случае каждый колебательный уровень расщепляется на четыре подуровня в соответствии с симметрией ячейки и получается четыре колебательных перехода. Картина смещений атомов из положений равновесия зависит от различия фаз колебаний отдельных пептидных групп. Японским физиком Миядзава была предложена теория возмущений с учетом взаимодействий слабо связанных между собой и достаточно локализованных пептидных колебаний. Частота колебания n ( d, d¢ )зависит как от степени взаимодействия, так и от разности фаз d между движениями атомов пептидных групп одной цепи и разности фаз d¢ колебаний пептидных групп через водородную связь. Таким образом, в нашем случае мы имеем четыре колебания: n (0, 0), n (0, p ), n ( p, 0 ), n ( p, p ), причем первое в ИК - спектре не активно.
С изучением спектров глобулярных белков дело обстоит сложнее. Регулярная структура полипептидной цепи в их молекулах образуется только на небольших участках, причем единое направление, вдоль которого могут быть ориентированы цепи, отсутствует, спектр получается намного сложнее.
1.6.23. Спектрополяриметрия
Метод дисперсии оптической активности широко применим при исследовании структуры и конформационных превращений в биополимерах, что объясняется сравнительной простотой измерений, отсутствием необходимости специального препарирования образцов в отличие от других методов, а также возможностью исследования малых концентраций.
Дисперсия оптической активности полипептидов и белков. Из уравнения
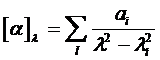 (1.6.36.)
(1.6.36.)
следует, что если известна сила вращения и длина волны каждой оптически активной полосы поглощения в данной молекуле, то можно предсказать дисперсию оптической активности. И наоборот, при помощи этого уравнения можно оценить электронные параметры соответствующих переходов из наблюдаемой дисперсии оптической активности.
Большинство оптически активных веществ не имеет полос поглощения в видимой части спектра: они расположены в ближней, а большинство - в далекой УФ - области спектра. Поэтому до тех пор, пока измерения проводятся в видимой и близкой УФ - области с не очень большой точностью, дисперсия оптической активности часто подчиняется одночленному уравнению Друде
![]() (1.6.37.)
(1.6.37.)
Хотя самая длинная полоса поглощения дает наибольший вклад, l с часто не совпадает со значением l длинноволновой полосы.
Характерная, т. е. не зависящая от белковых групп, но существенно зависящая от конформации полипептидов составляющая дисперсии оптического вращения возникает при взаимодействии белковых групп, входящих в состав полипептида. В a - спирали амидные группы ориетированы одна относительно другой вполне определенным образом. В результате между ними длинноволновая p®p*- полоса расщепляется на две с длинами волн l 1 и l 2 , в результате чего дисперсия оптического вращения спирального полипептида должна описываться двучленным уравнением Друде
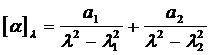 (1.6.38.)
(1.6.38.)
Т. к. расщепление невелико, то эту формулу можно подвергнуть преобразованию, в результате которого получается уравнение Моффита.![]()
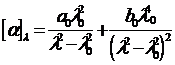 (1.6.39.)
(1.6.39.)
Из наклона прямой определяется b 0, а из пересечения с осью ординат - a 0. В этой формуле b 0 сильно зависит от выбора l 0.
В этом уравнении хорошо описывается дисперсия оптической активности спиральных полипептидов и фибриллярных белков.
Несмотря на значительные успехи, достигнутые в исследовании конформации макромолекул методами оптической активности, есть много неизученных вопросов, таких, как роль вторичной и третичной структуры белков и полипептидов.
1.6.24. Электрофоретические методы анализа белков.
Происхождение заряда белковой молекулы. Молекула белка приобретает заряд благодаря двум процессам: ионизации имеющихся ионогенных групп и связывания заряженных или способных к ионизации лигандов, включающих такие простые лиганды, как катионы и анионы буферных растворов. Однако основным источником заряда являются аминокислоты, среди которых наибольший вклад вносят аминодикарбоновые и диамино-карбоновые кислоты.
Из 10 ионогенных остатков аминокислот, входящих в полипептидную цепь, только три ( гистидин, лизин и аргинин )являются основными и придают молекуле белка положительный заряд; ряд группировок способны отдавать протон и придавать молекуле белка отрицательный заряд.
Влияние конформации белка на величину заряда определяется различными факторами, в частности большое значение имеет такое свойство белка, как
жесткость конформации.
В высокомолекулярном полиэлектролите, каким и является белок, суммарный заряд определяется главным образом свойствами экспонированных ионогенных групп, а ионогенные группы, ориентированные внутрь глобулы, вносят незначительный вклад. Влияние маскированных группировок становится все более заметным по мере денатурации белка.
При титровании число зарядов уменьшается и вместе с тем постепенно исчезает их влияние. Процесс титрования можно описать следующей серией реакций. Допустим, белок содержит сто групп, являющихся донорами протонов![]()
БН 100 ® БН![]() +Н+
+Н+
БН![]()
![]() БН
БН![]() +Н+
+Н+
. . . . . . . . . . . . . . .
БН![]()
![]() БН
БН![]() + Н+
+ Н+
БН представляет формулу белка, в которой связано n-протонов.
Если число диссоциированных групп вида i равно n, то константа будет соответствовать:
Кi = ![]() (1.6.40.)
(1.6.40.)
где aH+ - активность ионов водорода.
Рассматривая белковую глобулу как шар с равномерно распределенным поверхностным зарядом, можно доказать, что:
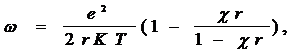 (1.6.41.)
(1.6.41.)
где r - радиус частицы, К - константа Больцмана, c - толщина ионной атмосферы, Т - абсолютная температура, w - электростатический фактор
( мера той дополнительной энергии, которая связана с процессом диссоциации ), который может быть использован для характеристики жесткости конформации.
Как правило, белки плохо растворимы в воде. В качестве дополнительного фактора стабилизации конформации выступают сорбированные ионы, которые придают глобуле дополнительный заряд и гидрофильность. Тот факт, что конфомационная стабильность белковой молекулы закодирована в аминокислотной последовательности, дает объяснение обратимости денатурации.
В реальных условиях реакция денатурации требует затраты энергии и осложняется постсинтетической модификацией, способной сильно изменить исходную структуру.
У денатурированного белка часть групп, обычно находящихся на поверхности глобулы, оказывается маскированной, и наоборот, ранее маскированные гидрофобные группы становятся экспонированными. Если в смеси имеется несколько фракций, то они обычно исчезают и появляется одна общая, т. е. имеет место электрофоретическая гомогенизация.
В целом рК аминокислотных остатков зависят от ближайшего окружения и бывают сдвинуты. Например, рК имидазола в разных белках варьирует от 6,4 до 9,4 и даже у одного белка группы одинакового типа, например фенольные группы хемотрипсиногена, часто имеют разные рК.
Фенольные и имидазольные группы имеют тенденцию показывать значение рК больше, чем ожидалось по модельным соединениям, а боковые аминогруппы - меньше. Другими словами, нативная структура благоприятствует существованию незаряженных фенольных и боковых аминогрупп, особенно в тех случаях, когда они находятся на мощном гидрофобном радикале ( лизин ).
Также надо учитывать, что заряд определяется для цельной белковой молекулы. Между тем, на большей поверхности белка существуют зоны только положительные или только отрицательные.
Из других факторов, определяющих рК групп, следует назвать: конформационные состояния, мало отличающиеся по свободной энергии и влияние водородных связей, которые в известной степени блокируют карбоксильные и аминные группы.

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 |
