
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
L. Mary-Rabine и соавт. (1978) регистрировали в ткани правого предсердия, резецированной у больных во время операции на сердце, ПД, образование которых замедлялось при добавлении адреналина в низкой концентрации и ускорялось при более высоких концентрациях адреналина. Эффект низких концентраций адреналина блокировался б-адреноблокатором фептоламином; эффект высоких концентраций — в-адреноблокатором пропранололом, что сходно с реакциями, происходящими в желудочковых волокнах Пуркипьо. Очевидно, что в мышце предсердий имеются б - и в-адренорецепторы.
В 1981 г. J. Loeb, J. Moran показали, что у собак при перфузии артерии СА узла ацетилхолииом или физостигмипом (ингибитор холинэстеразы) происходило демаскироваппе скрытых водителей ритма, которые оказались более чувствительными к парасимпатическим влияниям, чем СА узел. Последняя реакция может быть дополнительным механизмом, обеспечивающим главенствующее положение СА узла в предсердиях. Уровень автоматизма скрытых предсердных водителей ритма достигал 73% синусового автоматизма. Наиболее раннюю активацию предсердий при торможении СЛ узла авторы отметили вблизи заднего межузлового «тракта».
Группа авторов во главе с J. Воncau (1978, 1985) придерживается мнения, что множественные пред-сердпые водители ритма не являются полностью изолированными, а образуют вместе с СА узлом систему, названную ими «предсердным пейсмейкерным комплексом». В правом предсердии у собаки этот комплекс включает очаги автоматизма, которые распространяются в каудокраниальном направлении на протяжении 40— 50 мм, вблизи от пограничной борозды, между верхней и нижней полыми венами. Ширина зоны пейсмекеров составляет около 15 мм, а площадь — 45X15 (675 мм2), что в 10 раз больше размеров СА узла. Самые быстрые, краниальные, водители ритма располагаются позади rostrum СА узла в области Remak [Remak R., 1844], содержащей большое количество гап-глиозных клеток. Самые медленные, каудальные, водители ритма сосредоточены в зоне, известной как область Fredericq [Fredericq L., 1906]; она соответствует месту, где находятся предсердные водители ритма, продолжающие возбуждать умирающее сердце (ultium moriens). Особенность предсердного пейсмекерного комплекса состоит в том, что возбуждение может одновременно (или лишь с минимальным запаздыванием) начинаться по меньшей мере в двух центрах автоматизма. Благодаря такому взаимодействию обеспечивается быстрая деполяризация большого участка миокарда предсердий, например 900 мм2 за первые 10 мс. Как отдельные водители ритма, так и весь пейсмекерный комплекс регулируются парасимпатическими и симпатическими нервами [Boineau J. et al., 1985]. Недавно J. Boineau и соавт. (1988) представили данные о существовании предсердного пейсмекерного комплекса в сердце человека.
Автоматическая и триггерная активность выявляется в коронарном синусе, имеющем отношение к электрогемозу левого предсердия [Wil A., Cranei'ield P., 1977]. Кроме того, в левом предсердии обнаружены автоматические клетки вокруг устьев четырех легочных вен и в пучке Бахмапа.
В 1979 г. A. Wit и соавт. установили, что у человека и у экспериментальных животных мышечные клетки двустворчатого клапана обладают способностью спонтанно вырабатывать импульсы. По своей ультраструктуре эти клетки не отличаются от обычных предсердных клеток, однако им свойственна диастолическая деполяризация. Генерация импульсов опосредуется медленными каналами мембраны, поскольку она подавляется верапамилом и нечувствительна к ТТХ. При электрической стимуляции клеток и добавлении к ним адреналина либо при воздействии сердечных гликозидов появляются задержанные постдеполяризации и триггерная активность, устраняемая ацетилхолином и верапамилом. Таким образом, в створках митрального клапана могут при благоприятных условиях формироваться как медленные автоматические, так и более быстрые триггерные ритмы [, , 1981; Wit A. et al., 1979]. G. Rozanski и J. Jalife (1986) обнаружили, что такими же свойствами обладают мышечные клетки трехстворчатого клапана (в сердце кролика). Как митральный, так и трехстворчатый клапаны богато иннервированы парасимпатическими и симпатическими нервными волокнами. Адренергические и приходящие в определенное время холинергические стимулы способны увеличивать наклон диастолической деполяризации в клетках створчатых клапанов (парасимпатическое-симпатическое взаимодействие), т. е. усиливать их автоматизм.
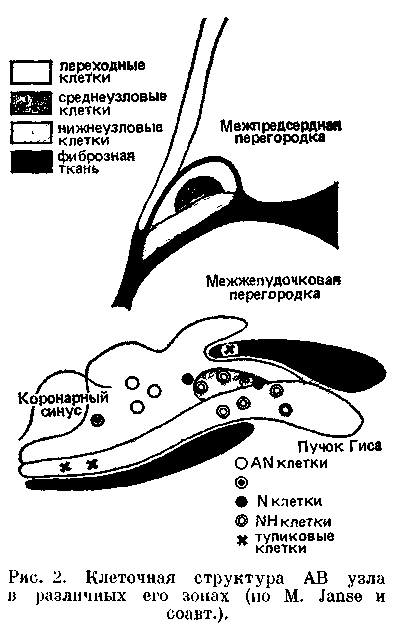
Атриовентрикулярный узел Ашоф-фа —Тавара [Tawara S., 1906; Aschoff L., 1908, 1910]. Он расположен в задней части межпредсердной перегородки, справа под эндокардом, впереди от устья коронарного синуса, непосредственно выше места прикрепления к перегородке септальнои створки трехстворчатого клапана; иначе говоря, — на правом треугольнике центрального фиброзного тела сердца [, , 1985; , 1987; Andersen R. et al., 1981; Bharati S. et al., 1983]. Оперирующие на проводящей системе сердца хирурги часто определяют положение АВ узла по треугольнику Коха [Koch W., 1912], в переднем углу которого находится АВ узел. Передневерхней стенкой треугольника Коха служит сухожилие Тодара (фиброзный пучок, проходящий в основании клапана нижней полой вены); нижнюю стенку образует место прикрепления к фиброзному кольцу септальнои створки трехстворчатого клапана; задневерхнюю стенку составляет устье коронарного синуса [Anderson R. et al., 1980].
При гистологическом и гистохими-ческом исследованиях выявляется трехслойное строение АВ уела, составленного из клеток, различающихся формой, размерами, контактами и, наконец, скоростью проведения импульса (рис. 2). Некоторые исследователи сравнивают структуру АВ узла с лабиринтом из специализированных волокон [Watanabe Y., Drei-fus L., 1965, 1980; Mazgalev Т. et al., 1986].
Проксимальный слой АВ узла, связанный с мышцей правого предсердия, представлен переходными (Т) клетками, удлиненными, по меньшими по величине, чем пред-сердные сократительные клетки. Группы переходных клеток отделены друг от друга прослойками кол-лагеновых волокон, резко замедляющих движение импульса. Н. Hecht и соавт. (1973) назвали этот слой преддверием АВ узла. Второй слой — собственно АВ узел, или компактный АВ узел ("Knotenpunkten" no S. Та-wara), имеет длину около 6 мм, ширину — 2—3 мм, толщину — 1,5 мм. Он составлен из плотно соприкасающихся клеток, среди которых, помимо переходных (Т) клеток, имеются округлые Р-клетки с малым числом органелл и миофибрилл. Их здесь заметно меньше, чем в GA узле [James Т., 1985].
Наконец, третий слой АВ узла — длинная дисталъная часть, непосредственно переходящая в атриовентри-кулярный пучок Гиса.
В свое время A. Paes de Carvalho (1959) выделил три электрофизиологические зоны АВ узла (в сердце кролика), отличавшиеся особенностями потенциалов действия: AN (atrium-nodus), N (nodus), NH (nodus-His). Теперь известно, что ПД типа AN вырабатываются в слое переходных клеток; ПД типа N — в небольшой группе клеток среднего, компактного, слоя и частично в переходном слое АВ узла; ПД типа NH — в нижнеузловом слое; в этом же слое найдены группы клеток, образующие так называемые электрофизиологические тупики, в которых угасает проведение импульсов (см. рис. 2).
Сложность строения АВ узла и других образований, связывающих предсердия с желудочками, многообразие их функций явились причиной появления нового термина—«АВ соединение». Правда, между морфологами и клиницистами существуют расхождения в определении этого понятия. Первые его толкуют расширительно, включая: 1) переходный слой и паранодальные волокна; 2) компактный АВ узел и его дистальную часть; 3) проникающий АВ пучок и 4) ветвящийся АВ пучок. Надо признать, что основания для такой трактовки дают некоторые эмбриологические данные. Однако, с клинической (электрокардиографической) точки зрения, к АВ соединению следует относить только первые три образования — до места разделения общего ствола на ветви. Соответственно проникающий АВ пучок входит в наджелудочковую область, а ветвящийся АВ пучок — в желудочковую. В последующем мы применяем термин «АВ соединение» именно в таком смысле.
У 80% мужчин и 93% женщин АВ узел снабжается кровью из ветви правой венечной артерии; у остальных людей кровь к АВ узлу поступает из огибающей ветви левой венечной артерии [ и др., 1982; James Т., 1961]. Важную коллатераль к АВ узлу образует артерия Кугеля [Kugel М., 1927]. АВ узел богато ин-нервирован парасимпатическими и симпатическими волокнами. Как и в СА узле, здесь имеется односторонняя избирательность иннервации: только левый блуждающий нерв вызывает отрицательный дромотропныи эффект в АВ узле; только левый симпатический перв укорачивает время АВ узлового проведения без воздействия на длину интервала Р—Р [Bal-sano F., 1981; Marigliano V., 1981; Randall W., Ardell J., 1985]. Наибольшая плотность нервных окончаний и рецепторов (в частности, мускарино-вых) отмечается в зоне N и в прилегающей к ней части зоны NH АВ узла [Mazgalew Т. et al., 1986]. На проведение импульса через АВ узел оказывают влияние вариации парасимпатического-симпатического взаимодействия, упоминавшегося выше [Ur-thaler F. et al., 1986]. В физиологических условиях существует параллельный регуляторный контроль частоты синусового ритма и АВ узловой проводимости, направленный на поддержание АВ проведения 1:1 [Вtrous G. et al., 1987].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 |
