
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Marriott (1966) назвал АВ узел «островом чудес в океане неизвестного». Это все еще актуальное определение не только отражает неполноту наших сведений об АВ узле, но и содержит признание важной роли, которую играет АВ узел в системе проведения и образования импульса. Основные функции АВ узла: 1) физиологическая задержка импульсов, движущихся от предсердий к желудочкам (синхронизация сокращений предсердий и желудочков с задержкой); 2) фильтрация (сортировка) предсердных волн возбуждения, препятствующая слишком частой активации желудочков; 3) защита желудочков от раннего, преждевременного возбуждения в уязвимой фазе их цикла.
О степени физиологической анте-роградной АВ задержки можно судить по следующим показателям: на преодоление импульсом небольшого АВ узла требуется от 50 до 130 мс (интервал А — Н Гис-электрограм-мы), т. е. намного больше времени, чем на прохождение длинного пути в правом предсердии. Еще медленнее осуществляется ретроградное вентрикуло-атриальное (ВА) узловое проведение (интервалы Н — А и V — А Гис-электрограммы). Например, у свиньи при стимуляции правого желудочка время ВА узлового проведения в среднем составляет 178±8,5 мс против 120± 18 для анте-роградного АВ узлового проведения [Bowman Т., Hughes N., 1984]. У 20% людей с неизмененным антероград-ным АВ проведением отсутствует ВА узловое ретроградное проведение [Mahmud R. et al., 1985]. Правда, во многих случаях ото связано с усиленным воздействием блуждающего нерва: после введения атропина ВА проведение восстанавливается. При ретроградном движении импульса через АВ узел прежде всего возбуждается нижнеперегородочная часть правого предсердия (иногда — область устья коронарного синуса — Gallagher J., 1987), затем все правое предсердие и с минимальным запаздыванием — левое предсердие.
Крайне низкая скорость распространения импульса в АВ узле (в среднем 0,05 м/с) зависит от преобладания в нем клеток с медленным электрическим ответом. В зоне N максимальный диастолический потенциал этих клеток равен — 60 мВ, небольшая пологая фаза О ПД устойчива к ТТХ, но блокируется ионами Мп++ и верапамилом. Между тем вопрос о месте наиболее выраженной задержки импульса в АВ узле все еще не вполне выяснен. Важным, хотя и не единственным, показателем скорости проведения импульса клетками является величина Vmax (dV/dt). По данным W. Tse (1986), Vmax в пара-подальных волокнах, подходящих к компактному АВ узлу, равняется 2,5 В/с, в самом АВ узле — 7 В/с, в стволе пучка Гиса — 30 В/с. Автор приходит к выводу, что АВ задержка волны возбуждения происходит в па-ранодальных волокнах у входа в АВ узел. Еще раньше М. Janse и соавт. (1976) указали, что от 20 до 80% времени А—Н и от 18 до 80% времени Н—А (Гис-электрограмма) приходится на прохождение зоны AN узла, что близко к данным W. Tse. Только 20% времени АВ проведения затрачивается на прохождение зоны N узла. Однако именно в этой небольшой зоне под влиянием усиливающейся вагусной стимуляции фрагментируется и утрачивает синхронность фронт возбуждения, что приводит к АВ узловым блокадам и, в частности, к развитию периодики Самойлова — Венкебаха.
Характеризуя АВ узел, следует коснуться еще двух его функций.
I из них имеет отношении к возможности продольного разделения АВ узла на два электрофизиологических канала — основу для re-entry и кругового движения импульса (рассмотрению этого механизма мы уделили место ниже — в разделах о реципрокных комплексах и АВ узловых реципрокных тахикардиях). Вторая функция имеет отношение к автоматизму АВ узла. В течение последних 30 лет допускалось, что автоматическая активность свойственна только клеткам NH-зопы АВ узла. Основанием для такого заключения послужили микроэлектродные исследования В. Hoffman и P. Granefied (1960), в которых не удалось in vitro обнаружить спонтанную диастолическую деполяризацию в клетках AN-и N-зоны АВ узла (в сердце кролика) . Однако позже некоторые авторы смогли зарегистрировать диастолическую деполяризацию в небольших изолированных препаратах АВ узла из сердца кролика [Kokuburn S. et al., 1980] и собаки [Tse W., 1986]. Недавно И. Ирисава и соавт. (1988) подтвердили в эксперименте, что клеткам АВ узла кролика присуща латентная пейсмекерная активность, однако в интактном сердце эти клетки никогда не возбуждаются спонтанно.
A. Wit и P. Cranefield (1982) так объясняют эти противоречивые данные. В здоровом (интактном) сердце кролика действительно отсутствует автоматическая активность в зонах AN и N АВ узла. Если же связь предсердий с АВ узлом нарушается, то в этих участках АВ узла можно уловить образование автоматических импульсов, частота которых превышает частоту синусового ритма, что исключает механизм сверхчастого подавления центров АВ узла. Причины угнетения автоматизма АВ узла представляются иными: поскольку предсердные клетки имеют более отрицательный потенциал покоя, чем клетки АВ узла, то возникающий между ними ток приобретает такое направление, которое предотвращает спонтанную диастолическую деполяризацию узловых клеток. Любой фактор, который ослабляет это элек-тротаническое межклеточное взаимодействие, будет способствовать проявлению скрытых автоматических потенций Л В узла (выраженный фиброз в место соединения правого предсердия и АВ узла и др. ' В 1983 г. R. Hariman и Chia-Maou С lien иришли к выводу, что у собак замещающий ритм АВ соединения может исходить из автоматических (Р) клеток зоны N АВ узла. Авторы напоминают о работе В. Scherlag и соавт. (1971), показавших, что оуаба-цн оказывает различное влияние на ритмы, берущие начало в АВ узле и стволе пучка Гиса.
Пока не вполне ясно, в какой степени эти экспериментальные данные приложимы к патологии человека, но изучение замещающих ритмов у больных с врожденными полными АВ блокадами подтверждает возможность возникновения проксимальных и дистальных АВ ритмов.
АВ пучок Гиса. Он служит продолжением дистальной части АВ узла [His W., 1893]. Начальный, или проникающий, сегмент пучка Гиса (общий ствол) длиной 10 мм проходит через центральное фиброзное тело в непосредственной близости от отверстий митрального и трехстворчатого клапанов и направляется вперед по верхнему краю мембранозной части мсжжолудочковой перегородки. Кол-лагеповые прослойки разделяют общий ствол на множество продольных пучков, слабо связанных друг с другом в поперечном направлении. Это обеспечивает разделение широкого фронта возбуждения на большое число равномерно движущихся изолированных волн, каждая из которых, по-видимому, достигает определенного предназначенного для нее участка миокарда желудков. Продольная диссоциация пучка Гиса, т. е. нарушение синхронности движения импульса, может имитировать изменения внутрижелудочковой проводимости [NarulaO., 1977, 1987].
Начинающийся у нижнего края мембранозной части межжелудочковой перегородки ветвящийся сегмент пучка Гиса представлен двумя ножками: правой и левой. Правая ножка образует изолированную ветвь длиной 4,5—5 см, направляющуюся вперед и вниз к различным субэндокар-диальным участкам правого желудочка и межжелудочковой перегородки. Первый функциональный контакт веточки правой ножки с сократительными волокнами происходит у основания передней сосочковой мышцы правого желудочка.
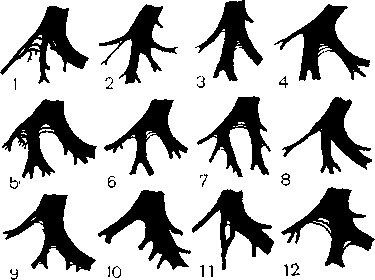
Левая ножка чаще начинается с широкого основания, лежащего суб-эндокардиально на левой стороне мышечной части межжелудочковой не-регородки. Направляясь слегка вперед и вниз, она вскоре разделяется на три главных разветвления, на что обратил внимание еще S. Tawara (рис. 3). В настоящее время большинство исследователей придерживаются мнения, что левая ножка (рис. 4) имеет мыоговар'иантное, трех-нучковое строение [Massing G. и James Т., 1976; Kulbertus Н. и Demou-lin J., 1977]. Тонкое и протяженное передневерхнее разветвление левой ножки (длина — 2—2,5 см, толщина — 3 мм) подходит к корню и середине передней сосочковой мышцы. Широкое и более короткое заднениж-нее разветвление (толщина — 6 мм), которое как бы служит продолжением общего основания ножки, распространяется к началу задней сосочковой мышцы. Центральное, или собственно переднее, разветвление левой ножки чаще берет свое начало из угла, образованного двумя другими разветвлениями, либо от одного из них. Оно направляется вниз к средней части межжолудочковой перегородки, поэтому его нередко называют среднеперегородочным (третьим) разветвлением левой ножки. Между ветвями левой ножки имеется широкая сеть анастомозов как в проксимальных, так и в дистальных участках [Lev M. et al., 1977].
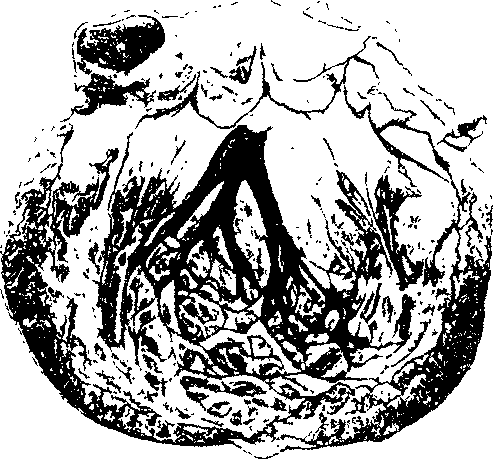
Рис.3. Фотография оригинального рисунка S. Tawara — распространение ветвей левой ножки в сердце человека [Tawara S. Des Reizlei-lungsystem der Saugo-tierherzens, 1906].
![]()

Рис. 4. Схема распространения волокон (ветвей) левой ножки в 20 сердцах людей,
не страдавших заболеваниями миокарда (по J. Demoulin, H. Kulbertus).
Основываясь на гистопатологических данных, полученных Y. Nakaya и соавт. (1987), D. Spodick (1988) называет проводящую систему левого желудочка многопучковой: «подобно вееру она широко распространяется по левой стороне межжелудочковой перегородки». При этом «двухпучковая концепция» М. Rosenbaum и соавт. (1970) сохраняет свое клинико-элек-трокардиографическое значение (см. гл. 16).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 |
