
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Реакция неизмененных и гипополяризованных волокон Пуркинье в ответ на одиночный преждевременный экстрастимул имеет общую направленность, однако в количественном отношении она неодинакова. В нормальных волокнах Пуркинье автоматизм временно прерывается с удлинением первого, возвратного, цикла на 30% по отношению к исходной длине цикла. В гипополяризованных волокнах возвратный цикл равен исходному циклу либо лишь слегка длиннее его. В части случаев преждевременные экстрастимулы, попадающие в определенную фазу автоматического цикла, могут вызвать аннигиляцию анормального водителя ритма, т. е. его устранение с внезапным прекращением выработки импульсов. В последующем функция анормального автоматического центра восстанавливается медленно и постепенно. «Аннигиляции» придается, с современной точки зрения, большое значение при объяснении интер-миттирования парасистолии [Jalife J., Anl/elevitchC., 1980].
Допускается, что анормальный автоматизм может быть причиной возникновения не только парасистолип, но и некоторых форм очаговых пред-сердньтх тахикардии, ускоренных ритмов. Возможно, что такой же механизм лежит в основе одной из разновидностей желудочковой тахикардии (ЖТ) в остром тгериоде инфаркта миокарда.
Это относится к очаговой автоматической ЖТ с частотой ритма ^200 в 1 мин, поскольку физиологический автоматизм волокон Пуркинье не может, как уже упоминалось, достигать столь высокого уровня [Crane-field Р., 1975].
Двумя процессами, названными ностдеполяризациями (afterdepolari-/ations), представлены формы нарушенного образования импульса, но связанные с автоматическими, т. е. самогенерирующими механизмами. Постдеполяризации — это вторичные подпороговые деполяризации (осцилляции мембранного потенциала), которые могут появляться: а) во время 2 и 3 фаз реполяризации ПД — их называют ранними постдеполяризациями; б) непосредственно после окончания ПД — их называют задержанными, или замедленными, постдеполяризациями [Cranefield P., 1975, 1977].
Ранние постдеполяризации. .Можно указать два важнейших условия их возникновения и связанных с ними трштерных ритмов. Первое условие: остановка или замедление реполяри-аации ПД, который начинается от достаточно большого потенциала покоя (между 75 и 90 мВ). Как показали В. Damiano и М. Rosen (1984), изучавшие влияние цезия хлорида на ПД волокон Пуркинье собаки, существуют два подвида ранних постдеполяризаций. Одни из них формируются при задержке реполяризации в фазе 2 ПД, т. е. на уровне мембранных потенциалов от —3 до —ЗОмВ. Другие появляются при задержке реполяризации в фазе 3 ПД, т. е. на уровне мембранных потенциалов от —50 до —70 мВ (рис. 9, 10). Напомним, что при таком же уровне потенциала мембраны может возникнуть анормальная спонтанная диастолическая деполяризация. Однако анормальный автоматизм—следствие частичной деполяризации мембраны, тогда как ранние иостдеполяризации — результат ее неполной реполяризации.
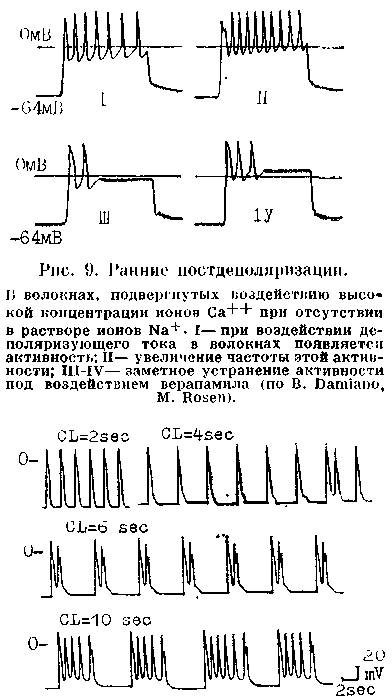
Рис. 10. Ранние постдеиоляризации. Влияние длины цикла стимуляции (CL) на вон иикшшоние ранних ностдшюляризаций и трштерного ритма: но море, удлинения цикла возрастает число ранних лостденоля-риуаций (но В. Damiano, M. Rosen).
Второе условие возникновения ранних постдеполяризаций и трштерных ритмов — урежепие основного ритма или частоты искусственной стимуляции. В эксперименте можно видеть, как при остановке реполяризации в фазе 2 или 3 ПД сначала регистрируются низкоамплитудные под-пороговые колебания мембранного потенциала, направленные кверху, т. е. в сторону более положительных потенциалов. Если частота основного ритма понижается, то происходит постепенное возрастание амплитуды ранних постдеполяризаций (в основном второго подвида). Достигнув порога возбуждения, одна из них вызывает образование нового ПД еще до окончания исходного ПД. Этот преждевременный ПД рассматривается
как триггерный, наведенный, ш>~ скольку он обязан своим возникновением ранней постдеполяризации, исходящей от основного ПД. В свою очередь, второй (наведенный) ПД может за счет своей ранней постдеполяризации вызвать третий, тоже триггерный ПД, а третий ПД стимулирует четвертый триггерный ПД и т. д. Следовательно, закрепляется пусковая ритмическая активность клеточной мембраны с различным числом импульсов. Надо учитывать, что при чрезмерном урежении основного ритма (или частоты стимуляции) амплитуда ранних постдеполяризаций начинает понижаться, и вероятность появления триггерного ритма уменьшается [Rosen M., 1986].
Триггерный ритм, вызванный ранними постдеполяризациями, не подвергается сверхчастому подавлению при коротком периоде стимуляции, но он может быть подавлен при более продолжительном периоде сверх частой стимуляция. Преждевременным экстрастимулом удается устранить такой триггерный ритм, если происходит гиперполяризация мембраны. В общем, триггерные ритмы этого типа исчезают в тот момент, когда по какой-либо причине полностью завершается процесс реполяризации, т. е. мембранный потенциал возвращается к своей максимальной физиологической величине (75—90 мВ).
Естественно, возникает вопрос о причинах, ведущих к задержке реполяризации, образованию ранних постдеполяризаций и триггерных ПД. Ответ мы находим в исследованиях D. Gadsby и P. Granefield (1977), обнаруживших два уровня мембранного потенциала, при которых волокнам Пуркинье свойственна устойчивая электрическая активность. Первый уровень равен —90 мВ, т. е. нормальному потенциалу покоя. Второй уровень соответствует —60 мВ, т. е. зоне триггерной активности. Остановка ре-поляризации на последнем уровне и образование ранних постдеполяризаций являются, согласно экспериментальным данным В. Damiano, M. Rosen (1984), характерным ответом клеток на различные факторы: гиперка-техолемию, гипокалиемию, ацидоз, гипокальциемию, ишемию, воздействие аконитина, N-ацетилновокаина-мида, соталола, цезия хлорида и др. Перерастяжение волокон Пуркинье при большом расширении или аневризме левого желудочка тоже создает условия для триггерных возбуждений.
Полагают, что следствием этих влияний бывает усиление входящего устойчивого деполяризующего тока, чувствительного к ТТХ (так называемого «тока Na+ окна») в фазе плато с удлинением ПД, вместе с торможением фазы 3 реполяризации, что, по-видимому, зависит от изменений тока ik, [Coulombe A. et al., 1985]. Еще раньше P. Cranefield (1977) показал, что колебания потенциала в фазах 2 и 3 реноляризации удается подавить верапамилом; в этом факте проявляется связь ранних постдеполяризаций с электрогенным Na—Са-обменным механизмом [Fischmeister R., Vas-sort G., 1981] и, возможно, с усилением входящего Са++-тока [January С. et al., 1988]. По последним данным, ранние постдеполяризации на уровне потенциалов от 0 до —30 мВ связаны с входящим Са++ током, переносимым через мембранные каналы L-типа [January С., Riddle J., 1989].
Что касается клинического значения ранних постдеполяризаций, то о нем можно судить по результатам исследований J. Brachmann и соавт. (1983). С помощью цезия хлорида они вызывали у собак синусовую бра-дикардию, удлинение интервала Q— Т и плеоморфную ЖТ. Авторы предполагают, что некоторые формы тахи-аритмий у больных с удлинением интервала Q—Т имеют триггерную природу. С. Mendez, M. Delmar (1985) подчеркивают роль этого механизма и формировании парасистолических очагов.
Задержанные постдеполяризации. Это электрические осцилляции в фазе 4 ПД, которым, как правило, предшествует гиперполяризация клеточ ной мембраны [Cranefield P., 1977]. Они изучены лучше, чем ранние постдеполяризации. Последовательность событий здесь такая же, как и при ранних постдеполяризациях. Подпо-роговое, демпфированное колебание мембранного потенциала себя не проявляет. Если же его амплитуда возрастает, достигая порога возбуждения, то возникает наведенный импульс—новый, преждевременный ПД (рис. 11). Этот ПД может, в свою очередь, быть источником другой пороговой осцилляции — ПД и т. п. В конечном счете формируется цепь триггер-ных возбуждений.
В эксперименте отмечено, что увеличение амплитуды задержанных ностдеполяризаций происходит тогда, когда в клетках повышается концентрация ионов Caf+. Медленный входящий Са++-ток не втянут непосредственно в этот процесс. Задержанные постдеполяризации генерируются «транзиторным входящим током», (iti), переносимым ионами Na+ и частично К+, но регулируемым внутриклеточной концентрацией ионов Са++, на которую влияет вхождение ионов Са++ в клетку [Marban E., Tsien R., 1982; Marban E., Smith Т., 1986; Gintant G., 1988].
Триггерную активность, вызываемую задержанными постдеполяризациями, изучали в волокнах Пуркинье, на которые воздействовали токсическими дозами сердечных гликозидов [Verrier G., 1977; Moak J., Rosen M., 1984], в предсердных клетках коронарного синуса, обработанных кате-холаминами [Wit A., Cranefield P., 1977; Wit A. et al., 1981], при экспериментальном инфаркте миокарда [Le Marek H. et al., 1985; Kimura S. et al., 1987], в предсердных и желудочковых волокнах человека in vitro [Dangman К. et al., 1982]. В 1988 г. S. Priori и соавт. впервые продемонстрировали in vivo (у кошек), что задержанные постдеполяризации можно вызвать путем раздражения левого звездчатого ганглия.
В отличие от ранних постдеполяризаций, возникновению (усилению) которых способствует брадикардия, задержанные постдеполяризации стимулируются учащением сердечного ритма. Это, по-видимому, происходит при синусовой тахикардии у больных с гипертрофией левого желудочка, кардиомиопатиями, ишемией миокарда. Вероятно, такой же характер носит «триггерный взрыв» — возникновение осцилляторной активности у некоторых больных вслед за периодом сверхчастой или программированной электрической стимуляции сердца [Wit A., Rosen M., 1983]. Весьма вероятно, что триггерную природу имеют ЖТ, вызываемые физической нагрузкой у больных без ИБС и других органических заболеваний сердца (см. стр. 309).

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 |
