
Партнерка на США и Канаду по недвижимости, выплаты в крипто
- 30% recurring commission
- Выплаты в USDT
- Вывод каждую неделю
- Комиссия до 5 лет за каждого referral
Хотя для возникновения спонтанной диастолической деполяризации п волокнах Пуркипье решающее значение имеет ток if, в этом процессе участвуют и другие ионные токи, в частности ток ik,, который и определяет зависимость автоматизма волокон Пуркинье от внеклеточной концентрации ионов К+, т. е. угнетение автоматизма при гиперкалиемии (этот ток незначителен в клетках СА узла, поскольку в них мало ik, каналов). В общем, в современной модели автоматизма волокон Пуркипье представлены четыре ионных механизма, зависящие от внеклеточной концентрации ионов К+: а) активация it, усиливающая пейсмекерную активность; б) активация ik, замедляющая или приостанавливающая пейсмекерную активность; в) активация тока Na—К насоса (ip), тоже замедляющая пейсмекерную активность; г) уменьшение тока ik, что усиливает пейсмекернуго активность.
В клетках СА узла складываются иные соотношения между ионными токами, определяющими их способность к автоматизму. На долю тока if приходится только до 20% этой активности. На передггий план выдвигается задержанный выходящий К+ ток (ik), угнетение которого во времени обеспечивает до 80% автоматической активности [Brown H. et al., 1982]. Разумеется, одними этими токами не ограничиваются сложные процессы, лежащие в основе спонтанного ритмического возбуждения клеток С А узла. Нельзя не упомянуть о токе ica, активация которого представляется необходимой для достижения порогового потенциала
Возбуждения этих клеток, Vmаx П амплитуды их ПД [Noble D., 1985]. Клиницистам хорошо известно, как чувствителен синусовый ритм к блокаторам Са каналов (L-типа) клеточной мембраны (верапамил) либо к B-адреноблокаторам (пропранолол), тоже влияющим на эти каналы через катехоламины. При анализе Са механизмов нельзя не учитывать активности Na+-Ka+-MeM6pairaoro насоса, Na+-Ca++-o6MeHnoro механизма, а также роли внутриклеточных систем, обеспечивающих секвестрацию и выделение ионов Са++. Накопление знаний в этой области биофизики происходит интенсивно, что позволит в ближайшем будущем еще больше приблизиться к истине [Noble D., 1985; Hagiwara N. et al., 1988; Levy M., 1990].
С электрофизиологических позиций, интервал меясду сокращениями сердца равен отрезку времени, п течение которого мембранный потепцп-ал покоя в клетках СА узла смещается до уровня порогового потенциала возбуждения. Три механизма оказывают влияние на продолжительность этого интервала и, следовательно, на частоту сердечной деятельности.
Первый из них (наиболее важный) — скорость (крутизна) диастолической деполяризации. При ее возрастании пороговый потенциал возбуждения достигается быстрее п происходит учащение синусового ритма (рис. 5). Противоположный эффект, т. е. замедление спонтанной диастолической деполяризации, ведет к урежению синусового ритма (рис. 6). Следует еще раз подчеркнуть, что С А узел не должен рассматриваться как однородный водитель ритма: в нем имеются группы клеток с различным уровнем автоматизма, и соответственно локализация пейсмекера в пределах СА узла может изменяться.
Второй механизм, оказывающий влияние тта уровень автоматизма СА узла, — изменение мембранного потенциала покоя его клеток (максимального диастолического потенциала). При увеличении этого потенциала (в абсолютных значениях), т. е. при гиперполяризации клеточной мембраны (например, под воздей ствием ацетилхолина), требуется больше времени для достижения порогового потенциала возбуждения, если, разумеется, скорость диастолической деполяризации остается неизменной. Следствием такого сдвига будет уменьшение числа сердечных сокращений в единицу времени.

Рис. 5. Ускорение спонтанной диастолической деполяризации в клетках Пуркинье под воздействием адреналина (no M. Otsuka),
I — до адреналина; П — через 10 мин после добанления адреналина.

Рис. 6. Влияние ацетилхолина на скорость спонтанной диастолической деполяризации в СА узле сердца кролика — замедление деполяризации с образованием длинных пауз (но М. Otsuka).
Третий механизм — изменение порогового потенциала возбуждения. Его смещение по направлению к нулю удлиняет путь диастолической деполяризации и способствует урежению синусового ритма. Приближение порогового потенциала к потенциалу покоя сопровождается учащением синусового ритма. Возможны и различные комбинации трех основных :>лектрофизиологиче-ских механизмов, регулирующих автоматизм СА узла. Соотношение между ними показано на рис. 7, 8.
Преобладание СА узла над остальными водителями ритма, рассеянными в проводящей системе сердца, обеспечивается прежде всего присущим ему более высоким уровнем автоматизма (скоростью спонтанной диастолической деполяризации), что приводит к опережающей разрядке латентных автоматических центров синусовыми импульсами. В этом проявляется «иерархия» автоматизма. Другой фактор, способствующий ведущей роли СА узла, — сверхчастое подавление (overdrive suppression — OS) скрытых пейсмекеров, т. е. временное подавление или угнетение их спонтанной активности после частой разрядки. В основе этого явления лежат уменьшение скорости спонтанной диастолической деполяризации, гиперполяризация автоматических меток и смещение их порогового потенциала возбуждения к менее отрицательным величинам за счет того, что эти клетки подвергаются активации, более частой, чем их собственная частота возбуждения. М. Vassalle (1977, 1985) показал, что торможение автоматизма при OS связано с тем, что Na+-K+-Hacocy мембраны требуется сравнительно много времени для удаления накопившихся в клетках ионов Na+.
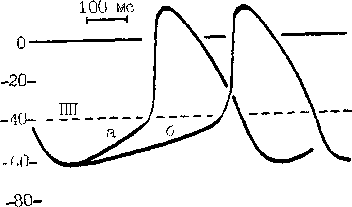
Рис. 7. Механизмы, изменяющие частоту синусового ритма.
Замедление синусового ритма при уменьшении скорости (крутизны) диастолической деполяризации (от а к б).
100 мс-80-
Рис. 8. Механизмы, изменяющие частоту синусового ритма. Замедление синусового ритма при смещении порогового потенциала (ПП) от ПП-1 до ПП-2 (от б к в); замедление синусового ритма при увеличении максимального диастолического потенциала от а до д (по В. Hoffman, P. Cranefield).
Еще один механизм, ограничивающий функцию скрытых водителей ритма, — электротоническое взаимодействие между сократительными и автоматическими клетками. В гл. 1 были представлены соображения A. Wit и P. Cranefield (1982) об электротоническом угнетении автоматизма клеток АВ узла предсердными клетками.
Изменения нормального автоматизма в СА узле или в скрытых автоматических центрах являются причиной возникновения таких аритмий, как: синусовые тахикардия, бради-кардия, аритмия, медленные (замещающие) и ускоренные выскальзывающие комплексы и ритмы (пред-сердные, атриовентрикулярные и идиовентрикулярные), миграция наджелудочкового водителя ритма, А В диссоциация.
АНОРМАЛЬНЫЙ АВТОМАТИЗМВпервые этот вид автоматической активности был обнаружен S. Imani-shi (1971) на препаратах волокон Пуркинье. Напомним, что физиологическая спонтанная диастолическая деполяризация в волокнах Пуркинт. р начинается от высокого уровня максимального диастолического потенциала (80—90 мВ) ("high potential automacity"). Она обеспечивает выработку за 1 мин 30—40 импульсов (при физиологической концентрации ионов К+) и возрастание импуль-сации до 100 в 1 мин под воздействием адреналина. Другая форма спонтанной диастолической деполяризации, названная анормальным автоматизмом, начинается в тех же клетках при их частичной деполяризации, т. е. снижении их максимального диастолического потенциала до 60— 40 мВ ("low potential automacity") [Cranefield P., 1975].
Такого рода явление можно наблюдать в препарате волокон Пуркинье собаки, обработанном бария хлоридом, а также в сохранивших жизнеспособность субэндокардиаль-ных волокнах Пуркинье сердца животного через 24 ч после перевязки венечной артерии и развития острого инфаркта миокарда [ и др., 1990; Dangman К., Hoffman В., 1980, 1985]. Спонтанно возникающим ПД присущ медленный электрический ответ, что связано с низким уровнем мембранного потенциала покоя поврежденных клеток Пуркинье. Частота же их автоматического ритма лыше физиологической, т. е. свойственной этим клеткам [Gilmonr R., 7л-pes D., 198-1].
Ряду авторов удавалось вызвать анормальную автоматическую активность в частично деполяризованных сократительных клетках предсердий и желудочков [Surawitz В., Imani-shi S., 1976; Katzung B. et al., 1977, 1985]. D. Singer и соавт. (1981) находили аналогичную по характеру активность в ткани предсердий, резецированной у больных, страдавших приступами очаговой (фокусной) ггредсердной тахикардии.
Рассматривая механизмы анормального автоматизма, В. Katzung и соавт. (1985), указывают на то, что его главным компонентом является зависящий от времени выходящий К+ ток ik. Однако имеются данные об участии в этом процессе и входящих в клетку Na+ и Са++ токов. В частности, R. Gilmour и D. Zipes (1984) подчеркивают сходство между ПД, генерируемыми частично деполяризованной клеточной мембраной, и ПД автоматических клеток СА п АВ узлов. Амплитуда тех и других, а также скорость диастолической деполяризации возрастают под влиянием катехоламинов и повышенной концентрации ионои Са++ в среде. Верапамил, дилтиазем и резкое снижение внеклеточной концентрации ионов Са++ могут полностью подавить анормальную автоматическую активность.
В отличие от физиологического автоматизма, анормальный автоматизм в гипополяризованных волокнах Пуркинье устойчив к сверхчастому подавлению, т. е. после его окончания патологические а втоматические импульсы появляются с прежней или даже с большей частотой. Возможно, что эти различия в быстроте выскальзывания нормальных и анормальных автоматических импульсов следует учитывать в клинике при дифференциальной диагностике двух типов автоматизма. Правда, если сверхчастая стимуляция длится больше 15 с, то автоматизм гиполя-ризованных волокон Пуркиньо тоже оказывается несколько подавленным IDangman К., Hoi'fman В., 198,4; Ro-son M., 1986, 1988].

|
Из за большого объема этот материал размещен на нескольких страницах:
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 |
